Title: Long-term litter decomposition controlled by manganese redox cycling
Download website:https://www.pnas.org/content/112/38/E5253
Significance
The rate-controlling mechanisms of litter decomposition are of fundamental importance for ecosystem nutrient cycling, productivity, and net carbon (C) balance. Current C cycling models rely primarily on climatic factors and lignin content as the main predictors of litter decomposition rates. Here, we show how the ability of the integrated plant–soil system to promote active redox cycling of manganese (Mn) regulates litter decomposition. Our work suggests that incorporating the coupling of litter decomposition and other elemental cycles, such as the Mn cycle, into conceptual and numerical models may significantly improve our mechanistic understanding and predictions of C cycling in terrestrial ecosystems.
Abstract:Litter decomposition is a keystone ecosystem process impacting nutrient cycling and productivity, soil properties, and the terrestrial carbon (C) balance, but the factors regulating decomposition rate are still poorly understood. Traditional models assume that the rate is controlled by litter quality, relying on parameters such as lignin content as predictors. However, a strong correlation has been observed between the manganese (Mn) content of litter and decomposition rates across a variety of forest ecosystems. Here, we show that long-term litter decomposition in forest ecosystems is tightly coupled to Mn redox cycling. Over 7 years of litter decomposition, microbial transformation of litter was paralleled by variations in Mn oxidation state and concentration. A detailed chemical imaging analysis of the litter revealed that fungi recruit and redistribute unreactive Mn2+ provided by fresh plant litter to produce oxidative Mn3+ species at sites of active decay, with Mn eventually accumulating as insoluble Mn3+/4+ oxides. Formation of reactive Mn3+ species coincided with the generation of aromatic oxidation products, providing direct proof of the previously posited role of Mn3+-based oxidizers in the breakdown of litter. Our results suggest that the litter-decomposing machinery at our coniferous forest site depends on the ability of plants and microbes to supply, accumulate, and regenerate short-lived Mn3+ species in the litter layer. This observation indicates that biogeochemical constraints on bioavailability, mobility, and reactivity of Mn in the plant–soil system may have a profound impact on litter decomposition rates.
Materials and Methods:we investigated the impact of biotic Mn redox cycling on long-term litter decomposition in forest ecosystems, taking advantage of a field experiment conducted in an old-growth Douglas-fir forest in the Oregon Cascades. During this 7-y experiment, each year’s litterfall was spatially separated by nylon mesh placed on the forest floor. The mesh created a series of six well-confined litter layers (a “littercake”) containing Douglas-fir needles at increasing decomposition stage. Our approach was to resolve Mn transformation and litter decomposition within the forest ecosystem at two different scales—across the whole soil profile and in microenvironments on needles colonized by fungi. Across the litter layer, we hypothesized that Mn oxidation covaries with the breakdown of aromatic litter components (i.e., lignins and tannins). Changes in Mn chemistry were identified by selective extractions and X-ray absorption near edge structure (XANES) spectroscopy, whereas the alterations in the molecular composition of litter was determined using a combination of Fourier-transform infrared (FTIR) spectroscopy, NMR, and laser desorption postionization mass spectrometry (LDPI-MS). Within microenvironments on needle surfaces, we anticipated that oxidative degradation of aromatic compounds occurs where fungi produce Mn3+. To spatially resolve Mn oxidation state and chemical transformations in these microenvironments, we applied a multimodal chemical imaging approach that coupled microscale X-ray fluorescence (µXRF)/X-ray absorption spectroscopy (XAS) with µFTIR imaging.
Results
Mn Transformations.
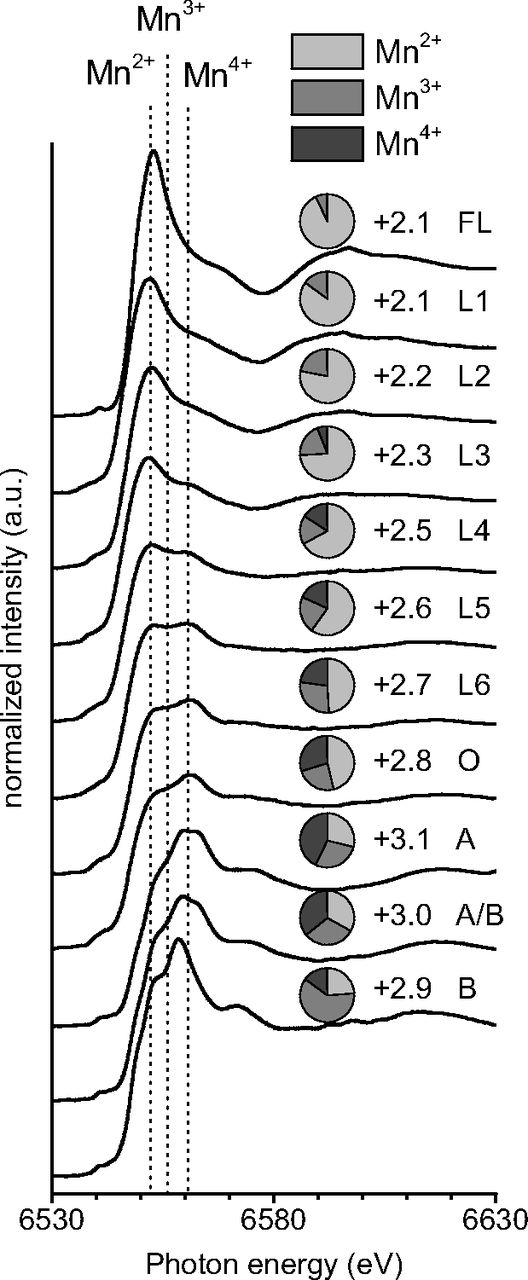
Fig. 1.
Change in Mn speciation in Douglas-fir needle litter (L) over 6 years of decomposition. Mn K-edge XANES spectra with approximate positions of absorption maxima for Mn2+, Mn3+, and Mn4+ forms. Pie chart Insets show the relative amounts of Mn2+, Mn3+, and Mn4+ with the adjacent number representing the average oxidation state as determined by linear combination fitting. Fresh needle litter (FL) and the underlying organic (O) and mineral (A and B) horizons were included for comparison.
Litter Decomposition.
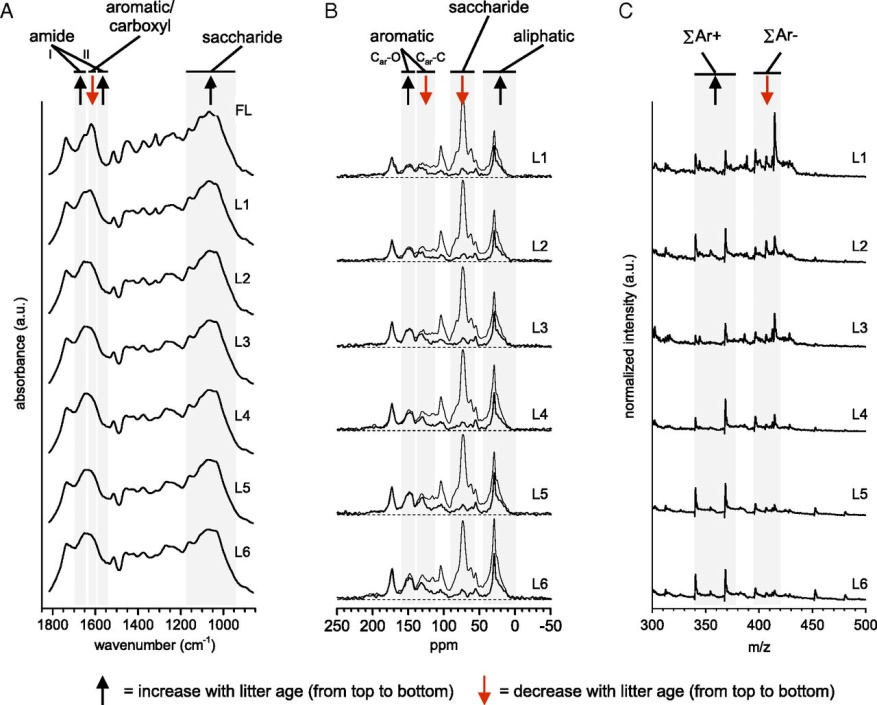
Fig. 2.
Chemical transformations of Douglas-fir needles over 6 y of decomposition. (A) FTIR spectra of needle litter. Absorbance of both (i) amide I and II groups associated with bacterial/fungal protein and chitin increased and (ii) C-O-C groups of polysaccharides increased with litter age whereas (iii) absorbance of COO-stretch of carboxylates and the ar-C-C stretch of aromatic lignin decreased. (B) 13C-NMR spectra of needle litter. Gray lines show spectra obtained by unselective CP/TOSS experiments including all carbons, whereas black lines show spectra obtained by selective CP/TOSS-DD sensitive to nonprotonated carbons and mobile carbons (e.g., -CH3 and long-chain -(CH2)n-). (C) Synchrotron-LDPI mass spectra of needle litter. Mass spectra presented here were collected with a photon energy of 9.5 eV and were chosen because they showed less fragmentation and greater signal-to-noise ratio.
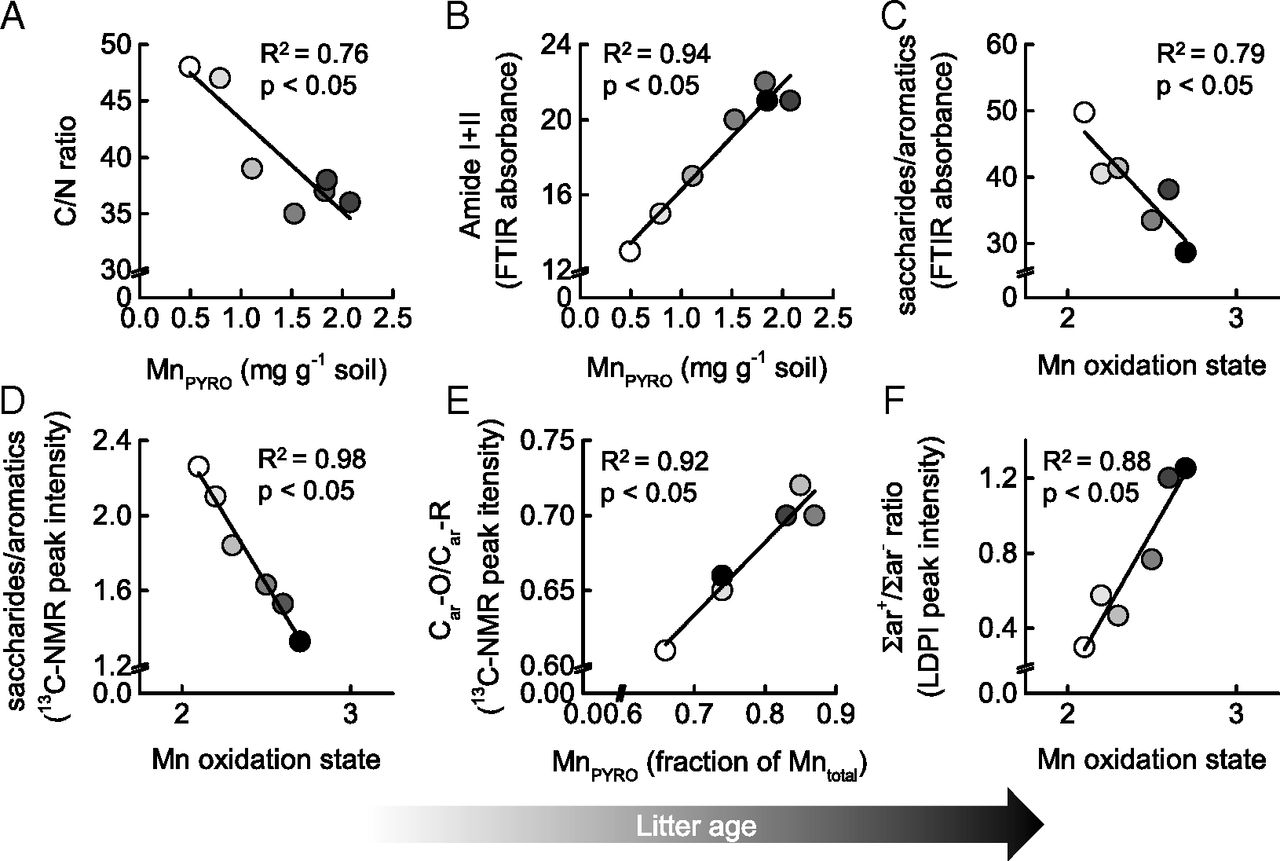
Fig. 3.
Covariation of litter and Mn transformations in Douglas-fir needles over 6 y of decomposition. (A) Changes in the C/N ratio across the litter layer in relation to pyrophosphate-extractable Mn (MnPYRO), here used as a proxy for bioavailable Mn. (B) Changes in the FTIR absorbance of amide I and II groups as a proxy for microbial protein/chitin abundance relative to variations in average Mn oxidation state. (C) Changes in litter decomposition (expressed by the ratio of saccharide/aromatic carbon absorbance in the FTIR spectra) in relation to Mn oxidation state. (D) Changes in litter decomposition (as indicated by the ratio of saccharide/aromatic carbon in 13C NMR spectra) in relation to Mn oxidation state. (E) Lignin transformations (expressed by the ratio of Car-O/Car-C in the 13C NMR spectra) in relation to average Mn oxidation state. (F) Changes in lignin transformations (expressed by the relative ratio in signal intensity of LDPI-detected aromatic mass fragments shown in Fig. 2C) in relation to average Mn oxidation state.
Mn Form and Distribution on Decomposing Needle Surfaces.
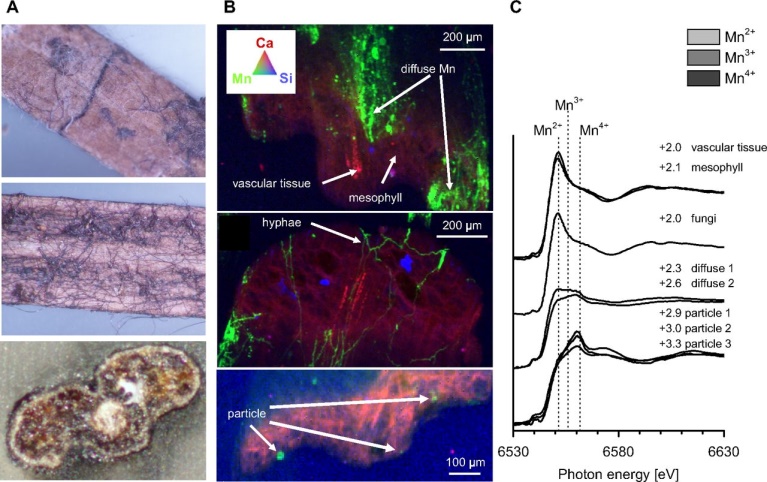
Fig. S1.
µ-XRF/XAS analysis of whole needle cross-sections taken from the top litter layer (year 1). (A) Photographs showing individual needles colonized by fungal hyphae associated with dark patches. (B) False color XRF maps of resin-embedded needle cross-sections showing the concentration of Mn (green) dense patches, hyphae and particles in contrast to Ca (blue) and Si (red). Color intensity is directly related to elemental concentrations. The apparent 3D appearance of the map is due to the penetration of X-rays ∼100–200 µm into the resin-embedded cross-sections. (C) Mn XANES spectra of the regions highlighted in B, illustrating the more oxidized state of Mn in diffuse patches and particles associated with the needle surfaces. Pie charts show the relative amounts of Mn2+, Mn3+, and Mn4+ and the number denoting the average valence state as determined by linear combination fitting .
Manganese Chemistry at the Hyphae–Epidermis Interface.
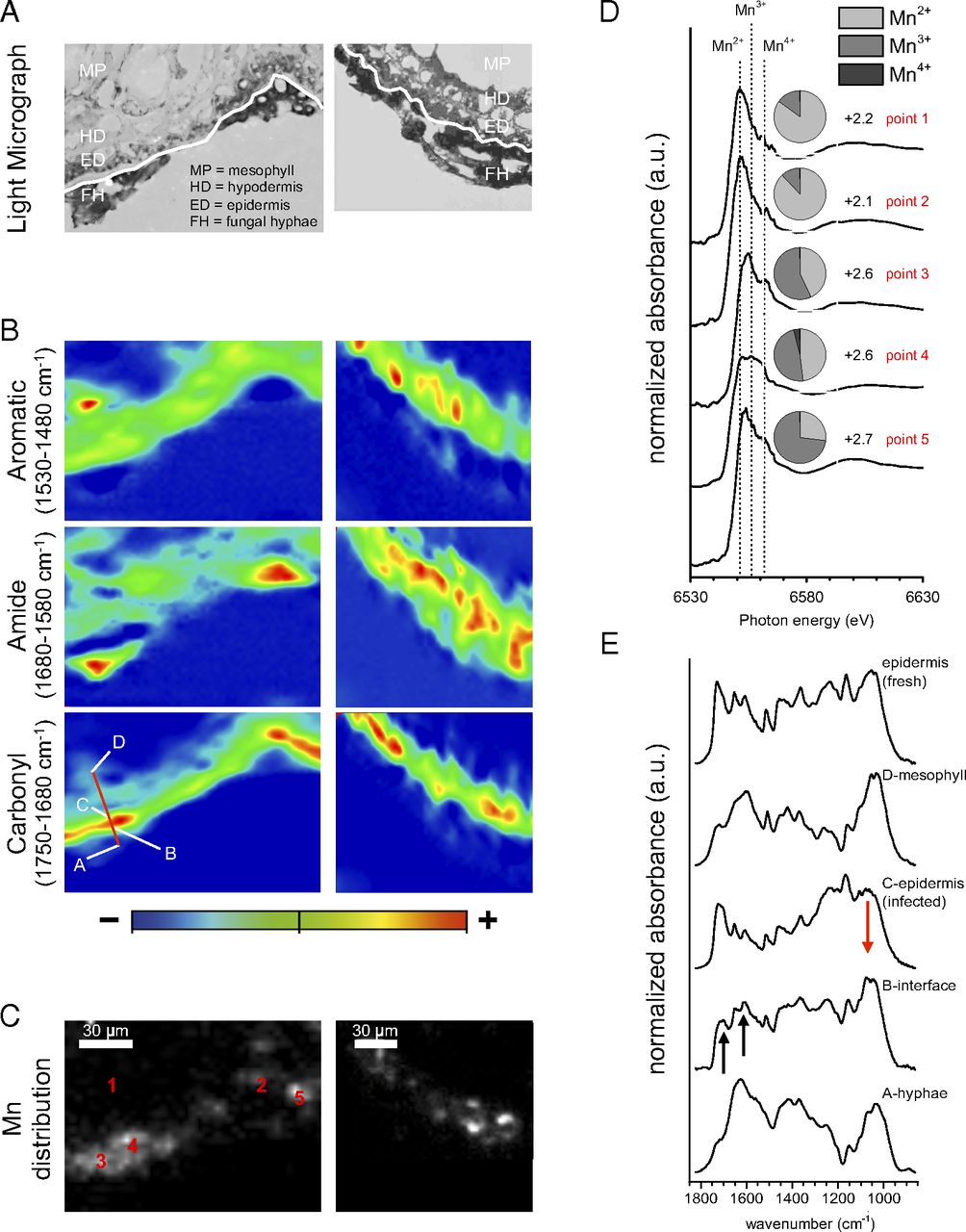
Fig. 4.
Mn transformations at the hyphae–epidermis interface of two decomposing needles. (A) Photographs of needle thin sections showing fungal hyphae (FH) colonizing the outer walls of epidermis cells (ED), partially fungal-infected hypodermis cells (HD), and mesophyll tissue (MP). The white line delineates the boundary between fungal hyphae and the needle epidermis. (B) Corresponding µFTIR heat maps showing the distribution of aromatic, amide, and carbonyl functional groups. Heat maps were generated using the integrated absorbance of spectral regions given in the figure. (C) Corresponding Mn distribution maps of the same region of interest generated by µXRF. (D) Mn XANES spectra collected at locations along the hyphae–epidermis interface (points 1 and 2 in C) and the needle’s mesophyll tissue (points 4 and 5). Pie charts show relative amounts of Mn2+, Mn3+, and Mn4+ at each location, with numbers indicating the average oxidation state. (E) µFTIR spectra extracted from transect across the hyphae epidermis interface, shown as red line in B). Note that absorbance at ∼1,700 and ∼1,610 cm−1 at the interface (B) increases relative to (C) infected epidermis regions and a fresh needle epidermis. (Scale bars: 30 µm.)
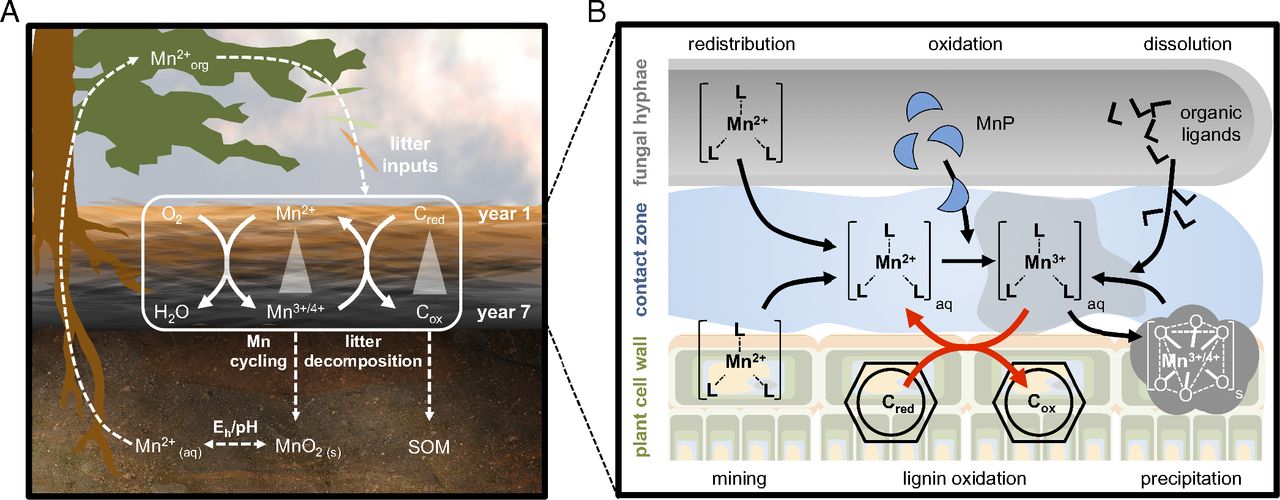
Fig. 5.
Macro- and microscale coupling of C and Mn cycles in forest ecosystems. (A) Mn cycling through the entire plant–soil system and its link to litter decomposition and SOM formation. (B) Microscale Mn cycling by litter-decomposing fungi and its role in the oxidative breakdown of lignin or other aromatic litter components.
Discussion
The results support the hypothesis that plant–soil systems in forest biomes have coevolved to optimize the “cell-wall degrading machinery”, thereby maximizing litter decomposition (and thus the recycling of nutrients) by ensuring the availability of key resources such as Mn (Fig. 5A). Because aspects of global change also impact ecosystem Mn fluxes, bioavailability in soils, plant uptake, and foliar litter concentrations , the tight coupling we demonstrate between Mn cycling and litter decomposition suggests that further research on regulators of ecosystem Mn fluxes is warranted.
By He Weihua