Title:Plant–fungal symbiosis affects litter decomposition during primary succession
https://doi.org/10.1111/oik.03648
Abstract:Microbial symbionts of plants can affect decomposition by altering the quality or quantity of host plant tissue (substrate) or the micro‐environment where decomposition occurs (conditioning). We studied the effect of the endophyte Epichloë sp. on litter decomposition in the Great Lakes dunes (USA) using a reciprocal design altering endophyte presence/absence in both American beachgrass Ammophila breviligulata substrate (litter bags) and its conditioning of the decomposition microenvironment. Symbiont treatments were crossed with rain‐out shelters that altered growing season precipitation. The first year of decomposition, senesced leaf substrate from A. breviligulata with Epichloë decomposed 21% faster than endophyte‐free substrate. By the third year, conditioning by live symbiotic plants reduced cumulative decomposition by 33% compared to plots planted with endophyte‐free plants. Of the traits we examined – litter quantity, C:N ratio, mineral composition, fungal colonization, and carbon chemistry – increased litter quantity via greater tiller production was the primary trait shift associated with endophyte symbiosis. Epichloë in A. breviligulata litter also altered litter nitrogen decomposition dynamics, as evidenced by lower nitrogen and protein content in decomposed tissue from plants that hosted the endophyte. Differences in initial litter quality and subsequent colonization by saprotrophic fungi were ruled out as key drivers. Altered precipitation had negligible effects on decomposing processes in the dunes. Grass–Epichloë symbiosis altered nutrient cycling through increasing the rate of litter decomposition when present in the litter and through reducing litter decomposition by conditioning the decomposition microenvironment. Epichloë are widespread symbionts of grasses. Thus, their effects on decomposition could be an important, but often overlooked, driver of nutrient cycling in grass‐dominated ecosystems.

Figure 1 Possible pathways for the effects of Epichloë on decomposition of host litter. The fungal endophyte may alter decomposition directly (solid lines) through the amount of litter production (substrate quantity) or the composition of the litter (substrate quality). The endophyte may also alter decomposition indirectly (dashed lines) by shifting the microenvironment in which decomposition occurs (conditioning). At the bottom of each box is the list of mechanisms that we examined for each pathway (in italics).
To assess the interactive effects of above‐ground plant symbionts and climate on decomposition in a native ecosystem, we manipulated Epichloë sp. in Ammophila breviligulata under alternative precipitation regimes. Specifically, we asked 1) does Epichloë affect decomposition directly by altering litter composition (substrate) or indirectly by altering the microenvironment for decomposition (conditioning)? 2) Does the precipitation regime directly affect decomposition or modify how the endophyte affects decomposition? 3) Do the traits that underlie changes in decomposition include shifts in litter substrate quantity, substrate quality, or colonization by saprotrophic fungi?
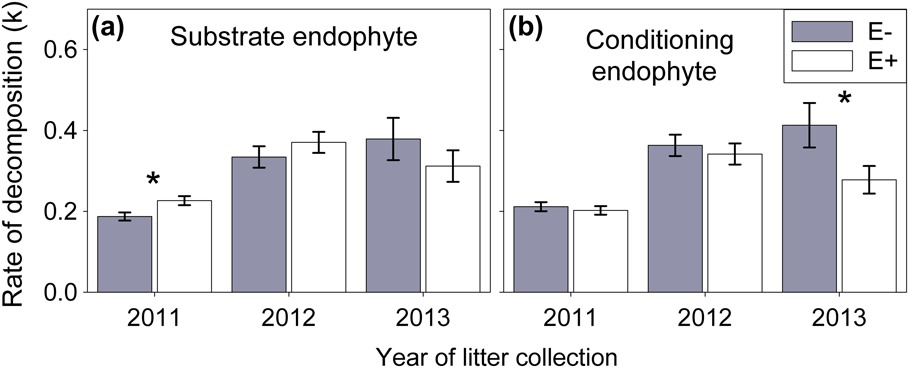
Figure 2 Rates of decomposition of Ammophila breviligulata substrate from litter bags deployed in May 2011 and collected in September 2011 (n=15), 2012 (n=15), or 2013 (n=14). (a) Epichloë (white bars (E+)) substrate decomposed faster than endophyte free substrate (filled bars (E−)) only during the first growing season (2011). (b) Litter bags (of either endophyte status) placed in plots conditioned by A. breviligulata with Epichloë (white bars (E+)) decomposed more slowly than litter bags placed in endophyte free plots (filled bars (E−)), after three growing seasons (2013). There was no interaction between substrate endophyte status and conditioning endophyte treatment.
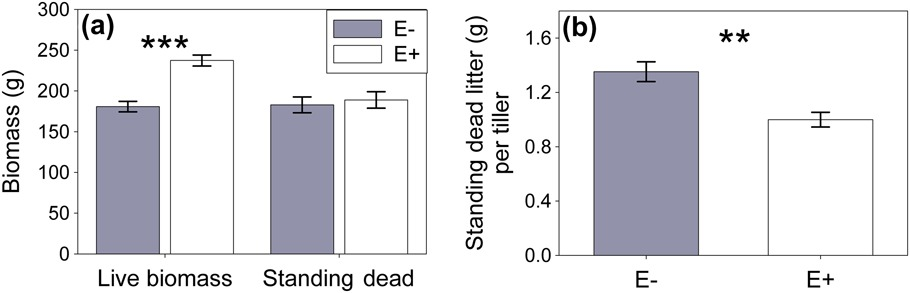
Figure 3 Ammophila breviligulata live biomass and standing dead litter per plot and per tiller (n=15). (a) Epichloë (white bars (E+)) increased the live biomass per plot for host plant A. breviligulata compared to endophyte free plots (filled bars (E−)); however, this increased biomass did not increase the amount of standing dead litter per plot. This was due (b) standing dead litter mass (g) per live tiller of A. breviligulata being lower in Epichloë plots than in endophyte free plots.
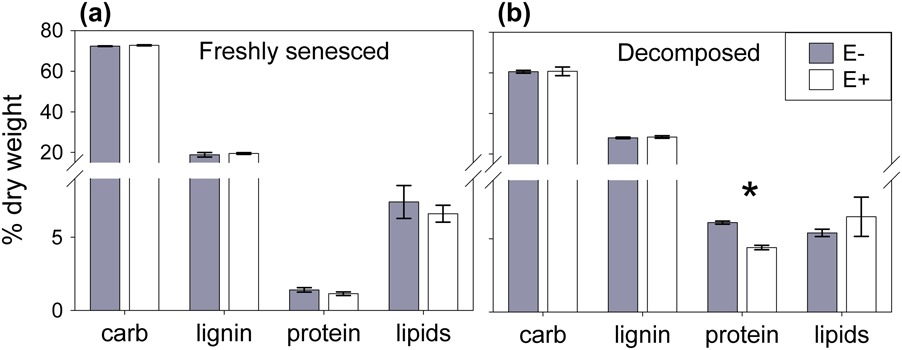
Figure 4 Carbon chemistry of Ammophila breviligulata litter, produced by plants with (white bars (E+)) or without (filled bars (E−)) Epichloë (n= 3). The presence of the endophyte did not affected percentage litter composed of carbohydrates (carb), lignin, protein, and lipids modeled from 13C NMR spectroscopy in (a) freshly senesced litter. However for (b) decomposed litter collected after three years, E+ substrate was depleted in protein compared to E− substrate.
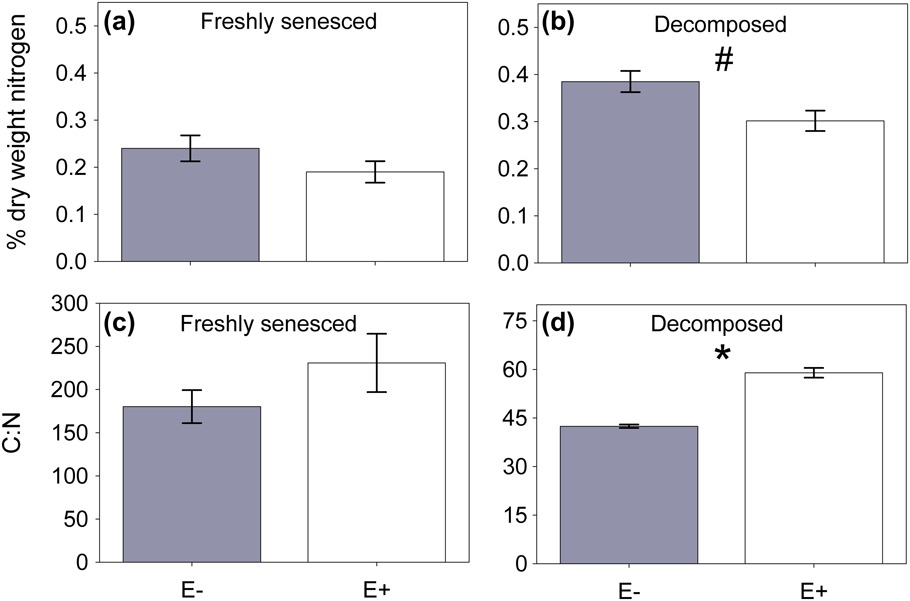
Figure 5 Percentage nitrogen and carbon:nitrogen (C:N) ratio of Ammophila breviligulata litter (n=3). Both (a) fresh and (b) decomposed substrate from A. breviligulata with the endophyte (white bars) tended to have lower nitrogen content than substrate from plants without the endophyte (filled bars). (c) There was no significant difference in C:N ratio between freshly senesced A.breviligulata substrate with Epichloë (E+) and endophyte free (E−) substrate. (d) The carbon:nitrogen ratio was significantly higher in bulked, decomposed substrate from plants with Epichloë than without Epichloë.
Discussion
Substrate produced by symbiotic plants had faster initial rates of decomposition than substrate from symbiont‐free plants. This increase in initial decomposition rate may have reduced the amount of standing dead litter per tiller in plots where the endophyte was present in live plants. However, the endophyte substrate treatment was not important after the first year of decomposition. Instead, the decomposition microenvironment (i.e. endophyte conditioning) became increasingly important, with endophyte presence in live plants in field plots reducing the rate of decomposition of both E+ and E– substrate types by the third year of decomposition. Mechanisms that may underlie these results include shifts in local nutrient availability, microbial activity, and microbial community composition, but are unlikely to be caused by initial differences in litter quality. These temporally dependent shifts in the importance of Epichloë in host plant litter decomposition could have large effects on the nutrient cycling in the nearly sterile dune soil.
Conclusion
This study is the first to report a temporal shift in the relative importance of the pathways through which above‐ground fungal symbionts can alter litter decomposition. Initial decomposition of A. breviligulata litter was faster if this substrate came from plants hosting the endophyte. This early increase in decomposition rate did not decrease the amount of standing dead biomass per m2 because the endophyte also increased aboveground plant biomass. Later in the decomposition process, endophyte symbiosis in living host plants slowed the rate of decomposition, perhaps by altering the soil microenvironment. Since previous research has shown that the endophyte is found ∼22% of A. breviligulata populations and its occurrence is spatially heterogeneous, both the effects of endophyte in the substrate and conditioning of the soil will likely lead to endophyte‐driven spatial heterogeneity in nutrient cycling within the dunes. This spatial heterogeneity may have important implications in successional processes as soil nutrients play an important role in plant succession. Given the widespread occurrence of Epichloë within grasses and more generally, of fungal symbionts in plants, temporal shifts in their influence on decomposition processes may have strong effects on nutrient cycling and carbon sequestration during succession in many ecosystems.