Article 1
Species-specific effects of soil fauna on fungal foraging and decomposition
DOI 10.1007/s00442-011-2005-1
Abstract: Decomposer fungi are primary decomposing agents in terrestrial soils. Their mycelial networks play an important role in nutrient mineralization and distribution, but are also nutritious resources for various soil invertebrates. Global climate change is predicted to alter the diversity and community composition of these soil fauna. To understand whether changes in invertebrate species diversity are likely to affect fungal-mediated decomposition, this study compared the grazing potentials of different invertebrate taxa and functional groups. Specifically, the grazing impacts of seven invertebrate taxa on the growth and spatial distribution of six basidiomycete fungi growing from beech wood blocks in soil microcosms were explored. Wood decay rates by fungi were also compared. The consequences of grazing were both taxon- and species-specific. Generally, macro-invertebrates caused the greatest damage, while meso- and micro-invertebrates often stimulated mycelial growth. Invertebrate size, preferences and population dynamics are likely to influence grazing potentials. Effects of grazing varied between fungi, with mycelial morphology and biochemistry possibly influencing susceptibility. Heavy grazing indirectly increased fungal-mediated wood decom- position. Changes in invertebrate community composition are predicted to have consequences for fungal growth, activity and community structure in woodland soils. Abiotic climate change factors including CO2 and temperature affect mycelial productivity directly, but the indirect effects, mediated through changes in the soil invertebrate community, may be equally important in controlling ecosystem functioning.
Results:
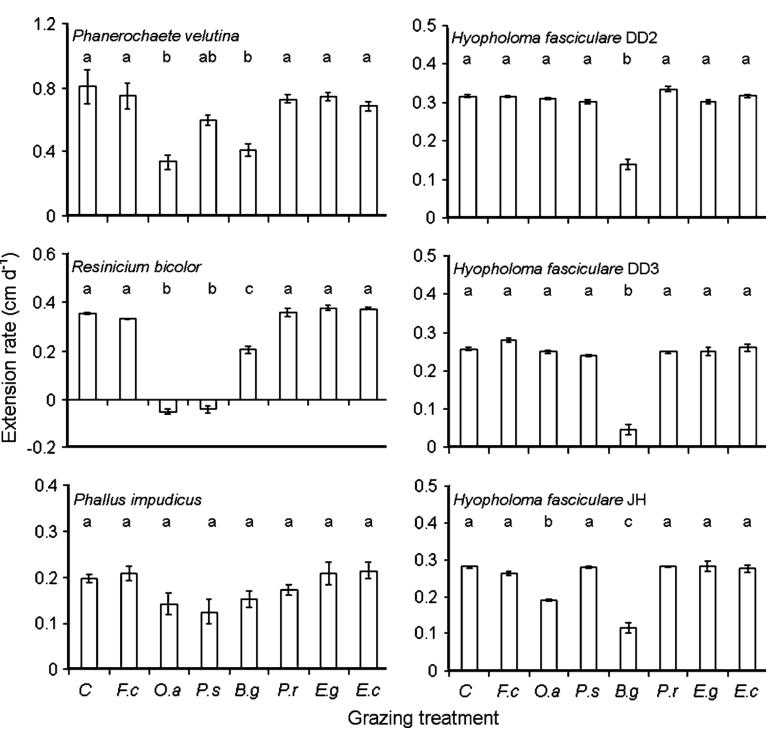
Fig. 1 Radial extension rates (mean ± standard error) of Phanerochaete velutina, Resinicium bicolor, Phallus impudicus, Hypholoma fasciculare DD2, Hypholoma fasciculare DD3, Hypholoma fasciculare JH growing across compressed non-sterile soil from a 2-cm3beech wood block during fungus-only control (C), Folsomia candida (F.c), Oniscus asellus (O.a), Porcellio scaber (P.s), Blaniulus guttulatus (B.g), Panagrellus redivivus (P.r), Euzetes globulus (E.g) or Enchytraeus crypticus (E.c) grazing treatments. Different letters above bars indicate significant differences in extension rates (ANCOVA; P ≤ 0.05); y axis scales vary between graphs
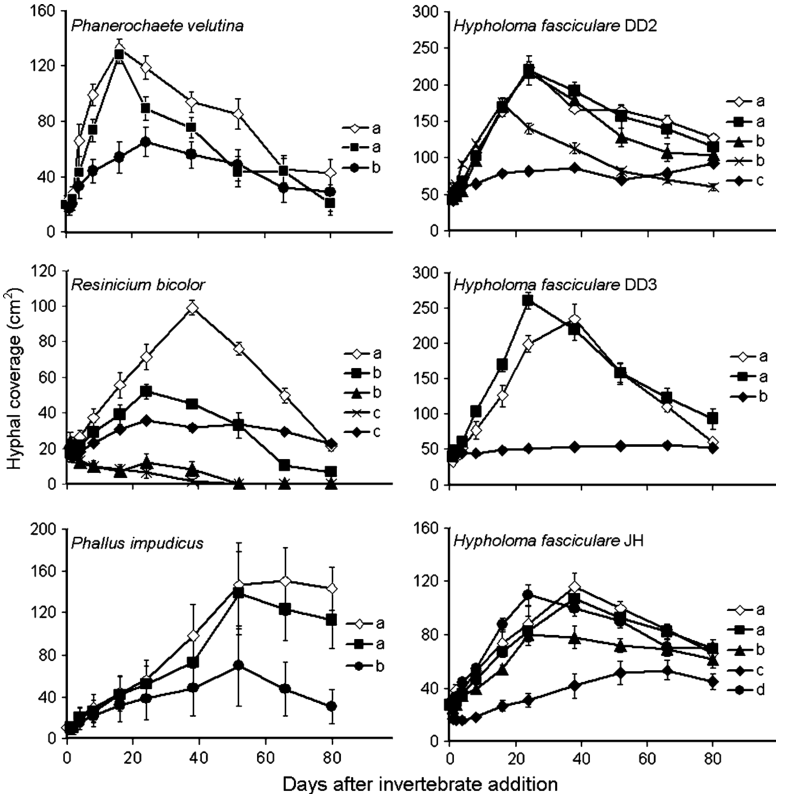
Fig. 2 Hyphal coverage of Phanerochaete velutina, Resinicium bicolor, Phallus impudicus, Hypholoma fasciculare DD2, Hypholoma fasciculare DD3, Hypholoma fasciculare JH, over 80 days during fungus-only control (open diamond), Folsomia candida (filled square), Oniscus asellus (filled triangle), Porcellio scaber (cross), Blaniulus guttulatus (filled diamond) or Panagrellus redivivus (filled circle) grazing treatments. Y axis scales vary between graphs. Different letters in the key indicate significant differences in hyphal coverage between treatments over time (repeated measures ANOVA; P ≤ 0.05). For clarity, not all grazing treatments are included; treatments which were not significantly different to control treatments were not shown
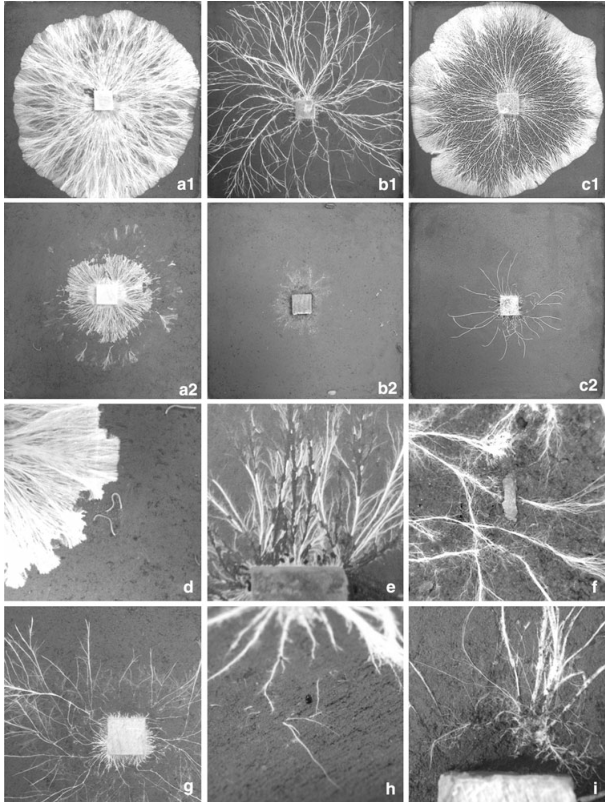
Fig. 3 Digital images showing un-grazed control growth of Hypholoma fasciculare DD2(a1), Resinicium bicolor (b1) and Phanerochaete velutina (c1), and grazing impacts of Blaniulus guttulatus (a2), Oniscus asellus (b2) and Folsomia candida (c2) respectively after 10 days of growth from 2*2*1 cm wood blocks on 24*24 cm soil trays. Also shows grazing styles of B. guttulatus on H. fasciculare (d), Porcellio scaber on R. bicolor (e), P. redivivus on P. velutina (f), Folsomia candida (g), Euzetes globulus (h) and Enchytraeus crypticus on R. bicolor (i)
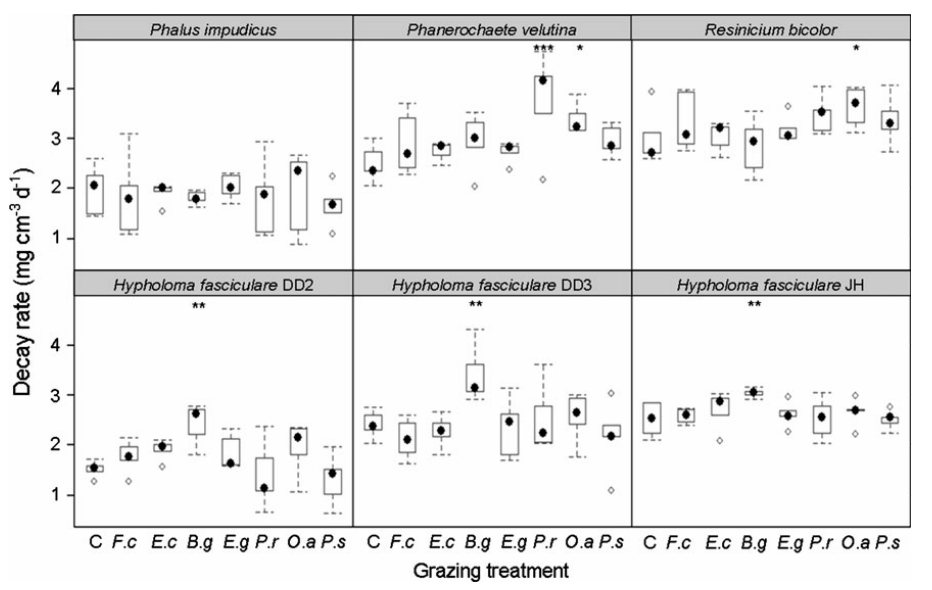
Fig. 4 Decay rates (mg cm-3) of beech (F. sylvatica) wood blocks colonised by Phallus impudicus (Pi), Phanerochaete velutina (Pv), Resinicium bicolor (Rb) and Hypholoma fasciculare strains DD2, DD3 and JH during control (C), Folsomia candida (F.c) Enchytraeus crypticus (E.c), Blaniulus guttulatus (B.g), Euzetes globulus (E.g), Panagrellus redivivus (P.r), Oniscus asellus (O.a) and Porcellio scaber (P.s) grazing treatments. Black dots indicate mean decay rates of five replicates of each treatment, boxes represent interquartile ranges and dotted lines indicate the range. Open circles represent treatment outliers. Stars indicate significant differences (ANOVA; P ≤ 0.05) from un-grazed controls (***P ˂ 0.001, **P ˂ 0.01, *P ˂ 0.05)
Invertebrate populations differentially affect mycelialgrowth, foraging and activity of basidiomycete fungi. Grazer-induced increases in wood decay rates further highlight the significance of these interactions for soil ecosystem functioning.
The present study suggests that predicted changes in invertebrate community composition (inter- and intra-specific) will also alter patterns of mycelial growth and functioning within soils. Changing invertebrate communities will also alter the selective pressures on fungal community compositions.
Conclusion:
Invertebrate populations differentially affect mycelial growth, foraging and activity of basidiomycete fungi. Selective grazing by invertebrate taxa could strongly influence fungal communities. Functional groups, determined by morphology, physiology and life-strategy, determine interaction consequences and a detailed characterization of these groups may provide compelling predictors of the effects of changing soil biodiversity on woodland decomposition (Gessner et al. 2010). Grazer-induced increases in wood decay rates further highlight the significance of these interactions for soil ecosystem functioning. While it is clear that climate change drivers such as elevated temperature or CO2affect fungal activity directly (Gange et al. 2007), the present study suggests that predicted changes in invertebrate community composition (inter- and intra-specific) (Jones et al. 1998; Wolters et al. 2000; Bokhorst et al. 2008) will also alter patterns of
mycelial growth and functioning within soils. Changing invertebrate communities will also alter the selective pressures on fungal community compositions (sensu Newell 1984). This has implications for nutrient cycling, carbon sequestration and productivity in soils under current and future climatic scenarios (Loreau et al. 2001; Wardle et al. 2004; H a ¨ttenschwiler et al. 2005).
Article 2:
Outcomes of fungal interactions are determined by soil invertebrate grazers
doi: 10.1111/j.1461-0248.2011.01682.x
Abstract: Saprotrophic fungal community composition, determined by the outcome of competitive mycelial interactions, is one of the many key factors affecting soil nutrient mineralization and decomposition rates. Fungal communities are not generally predicted to be regulated by top-down factors, such as predation, but rather by bottom-up factors, including resource availability. We show that invertebrate grazers can exert selective pressures on fungal decomposer communities in soil, reversing the outcomes of competitive interactions. By feeding selectively on the cord-forming fungus Resinicium bicolor, isopods prevented the competitive exclusion of Hypholoma fasciculare and Phanerochaete velutina in soil and wood. Nematode populations also reversed the outcomes of competitive interactions by stimulating growth of less competitive fungi. These represent two opposing mechanisms by which soil fauna may influence fungal community composition and diversity. Factors affecting soil invertebrate communities will have direct consequences for fungal-mediated nutrient cycling in woodland soils.
Hypotheses:
(i) selective grazing can reverse the outcomes of competitive fungal interactions with subsequent shifts in fungal species composition; (ii) as a result of increased grazing intensity, macrofauna will exert greater selective pressures on fungal communities than meso and microfauna; and (iii) grazing will influence fungal-mediated wood decay.
Results:
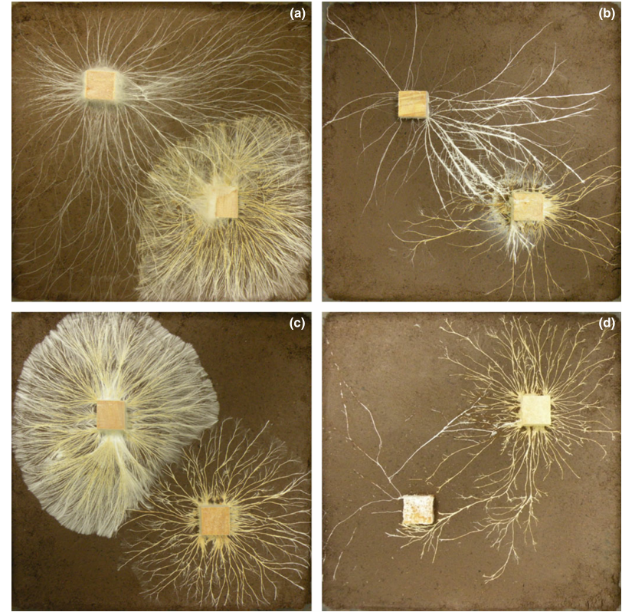
Figure 1 Digital images showing examples of the four possible outcomes of mycelial interactions in soil: overgrowth (a) of Hypholoma fasciculare DD3 (right) by Phanerochaete velutina (left), replacement (b) of H. fasciculare JH (right) by Resinicium bicolor (left), deadlock (c) between H. fasciculare JH (right) and DD3 (left), and mutual replacement (d) between H. fasciculare JH (right) and Resinicium bicolor (left).
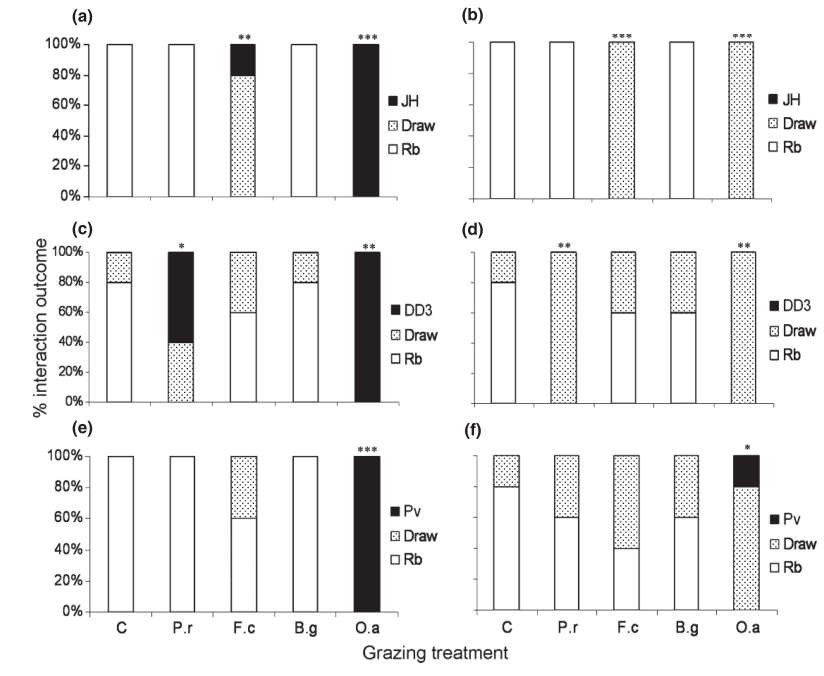
Figure 2 Percentage outcomes of competitive fungal interactions with Resinicium bicolor (Rb) against Hypholoma fasciculare JH (Hf JH), Hypholoma fasciculare DD3 (Hf DD3) and Phanerochaete velutina (Pv) in soil (a, c, e) and wood blocks (b, d, f) during control (C), Folsomia candida (Fc) Oniscus asellus (Oa), Blaniulus guttulatus (Bg) and Panagrellus redivivus (Pr) grazing treatments. Stars indicate significant differences (logistic regression) compared to ungrazed controls (***P ˂ 0.001, **P ˂ 0.01, *P ˂ 0.05). All other fungal interactions are not included, as P. velutina out competed both H. fasciculare strains in 100% of each grazing treatment and every interaction between the two H. fasciculare strains resulted in a draw.
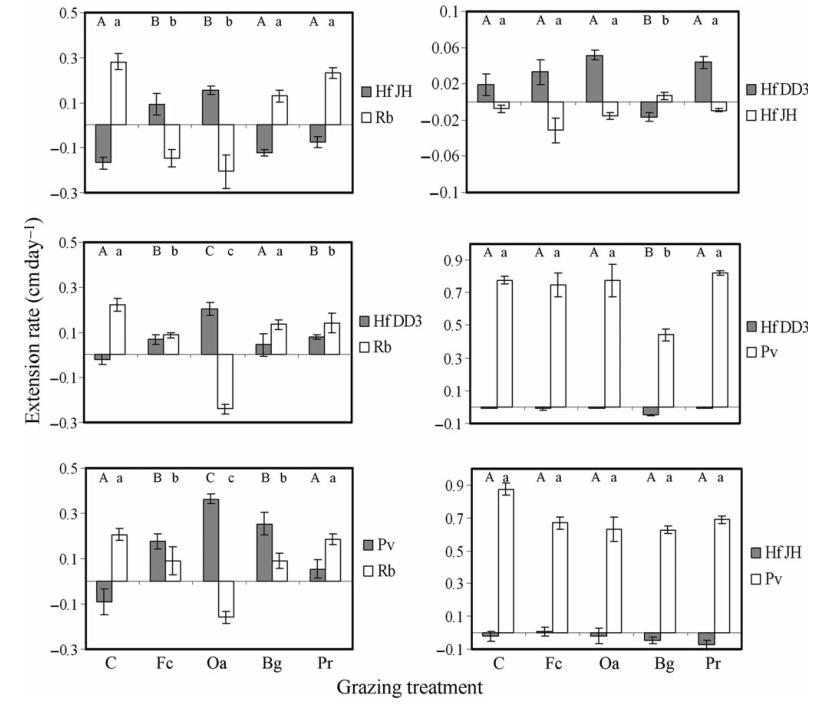
Figure 3 Extension rates of Resinicium bicolor (Rb), Phanerochaete velutina (Pv), Hypholoma fasciculare DD3 (Hf DD3) and Hypholoma fasciculare JH (Hf JH) growing towards one another in control (C), Folsomia candida (Fc), Oniscus asellus (Oa), Blaniulus guttulatus (Bg) and Panagrellus redivivus (Pr) grazing treatments. Negative values indicate that mycelia are retreating due to grazing or competitive interactions. Different letters indicate significantly (P ˂ 0.05; ANCOVA; Minitab 15) different extension rates. Upper and lower case letters refer to different fungi and were analysed separately. Y-axis scales vary between graphs.
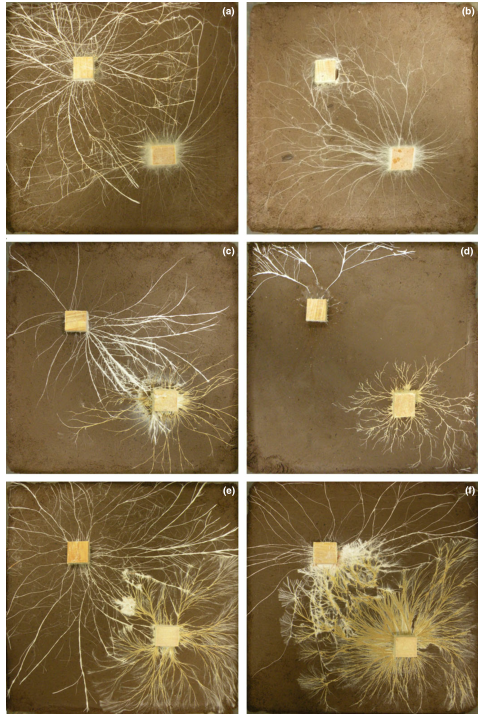
Figure 4 Digital images showing the outcomes of mycelial interactions with Resinicium bicolor against Phanerochaete velutina (a, b), Hypholoma fasciculare JH (c, d) and Hypholoma fasciculare DD3 (e, f) in ungrazed control (a, c, e) and grazed [Oniscus asellus (b), Folsomia candida (d), Panagrellus redivivus (f)] treatments. Competing mycelia extended from 2*2*1 cm wood blocks across 24*24 cm soil trays.
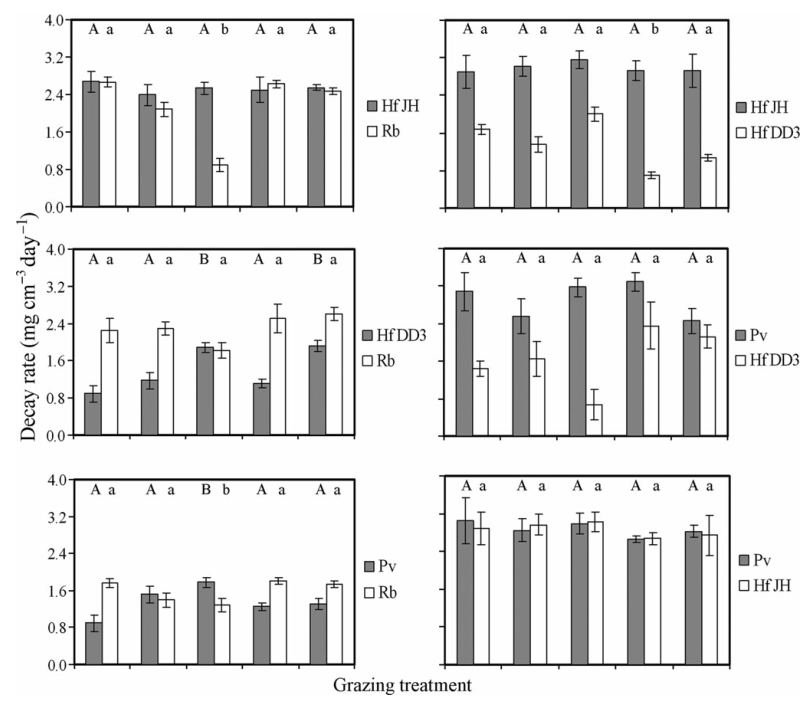
Figure 5 Decay rates of wood blocks colonised originally by Resinicium bicolor (Rb) against Hypholoma fasciculare JH (Hf JH), Hypholoma fasciculare DD3 (Hf DD3) and Phanerochaete velutina (Pv) during competitive mycelial interactions against one another in control (C), Folsomia candida (Fc), Oniscus asellus (Oa), Blaniulus guttulatus (Bg) and Panagrellus redivivus (Pr) grazing treatments. Different letters indicate significantly (P ≤ 0.05; two-way ANOVA; Minitab 15) different decay rates. Upper and lower case letters refer to different fungi and were analysed separately.
In the absence of grazing, P. velutina replaced H. fasciculare DD3 and JH, whereas all the three were replaced by R. bicolor.
Decay rates were significantly (F2,12= 10.22, P = 0.003) faster during interactions with H. fasciculare JH and DD3 than when competing with R. bicolor. Decay rates of wood blocks colonised by the three remaining fungi were not significantly (P > 0.05) influenced by any opposing fungal strain.
The presence of grazers affected the rate of decay of wood blocks colonised by P. velutina,R. bicolor and H. fasciculare DD3 (Fig. 5)
Significant (two-way ANOVA; fungus · invertebrate interaction: P ≤ 0.05) interactive effects of competing fungus and grazing treatments suggested that fungal opponents determined the potential of grazers to affect wood decay rates. Overall decay rates were, however, positively correlated (r = 0.521, n = 300, P < 0.001) with mycelial extension rates of the colonising fungi. This suggests that any reduction in wood decay was the result of reduced mycelial growth, following intensive grazing or replacement by an opponent.
B. guttulatus reduced H. fasciculare DD3 wood decay rates during intraspecific interactions with H. fasciculare JH (F6,68= 11.93, P = 0.01; Fig. 5). This is the only confirmed direct influence of grazers on decay rate; as neither opponent reached the opposing wood block, the decreased decay of H. fasciculare DD3 was the direct result of reduced fungal biomass during B. guttulatus grazing. Grazers can determine fungal dominance via two opposing mechanisms: (1) restriction of the more competitive fungus or (2) stimulation of the less competitive species. Phanerochaete velutina, for example, produces more cellulytic enzymes (Crowther et al. 2011c), and decomposes wood at a faster rate (Tordoff et al. 2008) than R. bicolor when growing alone. The potential of O. asellus to shift fungal communities in favour of the former may therefore lead to increased nutrient mineralisation and wood decomposition rates.
The aforementioned changes in fungal community structure are likely to influence litter decomposition rates (i.e. if a slow decomposer fungus is replaced by a more rapid species, overall decay rates will be increased).
For example, decay rates by R. bicolor were reduced during extra-resource mycelial grazing by O. asellus. This effect has been recorded previously, but in the present study, the removal of R. bicolor also stimulated mycelial growth and decomposition rates by P. velutina. Resulting decay rates were increased not only via the change in fungal community composition but also by the stimulated growth and activity of the promoted fungal species during grazing.
In conclusion, selective grazing by soil fauna will affect rates of nutrient turnover, both by altering fungal species composition, and also by modifying the relative capacities of interacting fungi to decompose wood
Article:Biotic interactions mediate soil microbial feedbacks to climate change
https://www.pnas.org/content/112/22/7033
Decomposition of organic material by soil microbes generates an annual global release of 50–75 Pg carbon to the atmosphere, ∼7.5–9 times that of anthropogenic emissions worldwide. This process is sensitive to global change factors, which can drive carbon cycle– climate feedbacks with the potential to enhance atmospheric warming. Although the effects of interacting global change factors on soil microbial activity have been a widespread ecological focus, the regulatory effects of interspecific interactions are rarely considered in climate feedback studies. We explore the potential of soil animals to mediate microbial responses to warming and nitrogen enrichment within a long-term, field-based global change study. The combination of global change factors alleviated the bottom-up limitations on fungal growth, stimulating enzyme production and decomposition rates in the absence of soil animals. However, increased fungal biomass also stimulated consumption rates by soil invertebrates, restoring microbial process rates to levels observed under ambient conditions. Our results support the contemporary theory that top-down control in soil food webs is apparent only in the absence of bottom-up limitation. As such, when global change factors alleviate the bottom-up limitations on microbial activity, top-down control becomes an increasingly important regulatory force with the capacity to dampen the strength of positive carbon cycle–climate feedbacks.
Significance:
The land carbon–climate feedback is incorporated into the earth system models that inform current Intergovernmental Panel on Climate Change projections. This feedback is driven by increases in soil microbial decomposition and carbon loss from soils under global change scenarios. The present study shows how trophic interactions in soil can mediate microbial responses to combined global change factors. As soil nitrogen deposition increases, the limitations on fungal growth are alleviated, stimulating total enzyme activity and decomposition rates. However, this process also affects the grazing activity of soil invertebrates. In the absence of nutrient limitation, top-down control by grazing isopods emerges as a dominant control, limiting any increases in fungal activity and carbon cycling.
Results:

Fig. 1. Digital images showing (A) the Harvard Forest warming plots (although the study was conducted in the fall, this image was captured in winter to delineate the warming plots clearly); (B) the arrangement of biotic treatment mesocosms within each abiotic plot; (C) the surface of the soil (under the litter layer) within a warming+nitrogen addition plot in the absence of isopods; and (D) the surface of the soil within a warming+nitrogen addition plot in the presence of grazing isopods.
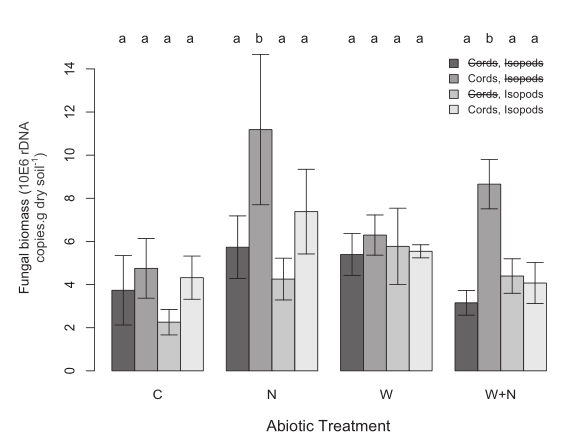
Fig. 2. Relative fungal sequence copies (mean ± SE) in treatment chambers, estimated using quantitative PCR. Fungal copy numbers were estimated by amplifying the ITS1/qITS2* universal primers. Bar color represents the biotic manipulation (darkest = −fungal cords,−isopods; second darkest = +fungal cords,−isopods; third darkest = −fungal cords,+isopods; lightest = +fungal cords,+isopods) within abiotic treatments (C, control; N, nitrogen addition; W, warmed; W+N, warming+nitrogen addition), showing how the presence of fungal cords enhanced fungal biomass in soil with nitrogen amendment, an effect that was mitigated by isopod grazing. Letters above the bars indicate significant (P < 0.05) differences between treatments. Note that error bars are included, but they cannot accurately represent the variation explained by the random effects of plot and soil moisture in the mixed-effects model.
Absent Macro-invertebrates:
Because of the overriding positive effect of soil nitrogen addition, fungal biomass in plots containing fungal cords was 1.8 times higher in future conditions (i.e., warming+ nitrogen addition plots) than in ambient controls.
Top-Down Control:
The inclusion of isopods in the +fungal cords , + isopods chambers prevented the climate-induced increases in fungal biomass.
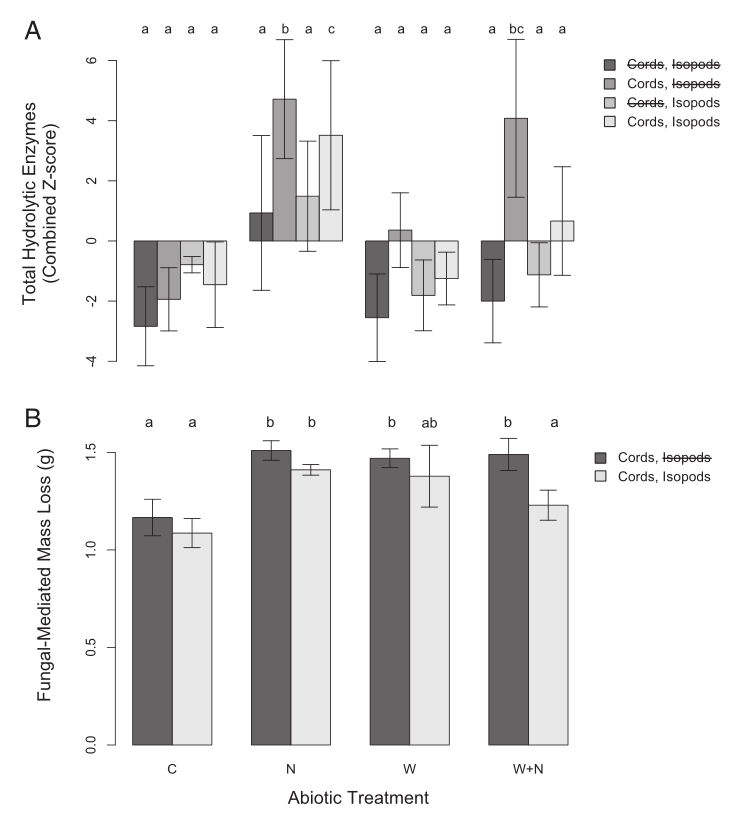
Fig. 3. Total standardized hydrolytic enzyme production (A) and wood decomposition (B) rates across all treatments. Z-scores were calculated for all enzymes to minimize the overriding effects of the most active enzymes detected. This value represents the number of SDs from the mean value for that enzyme across all 96 treatment chambers. Standardized values for each enzyme were summed to get an estimate of relative changes in total enzyme production. Values represent means across biotic and abiotic treatments (C, control; N, nitrogen addition; W, warmed; W+N, warming+nitrogen addition). Letters above bars indicate significant (P < 0.05) differences between treatments. Error bars cannot accurately represent the variation explained by the random effects of plot and soil moisture in our mixed effects model. Despite the effects of the global change factors on hydrolytic enzymes, no abiotic treatments significantly influenced the abundance of oxidative enzymes in soil, which were affected only by the presence of cord-forming fungi (F1,53= 17.740891, P = 0.0001).
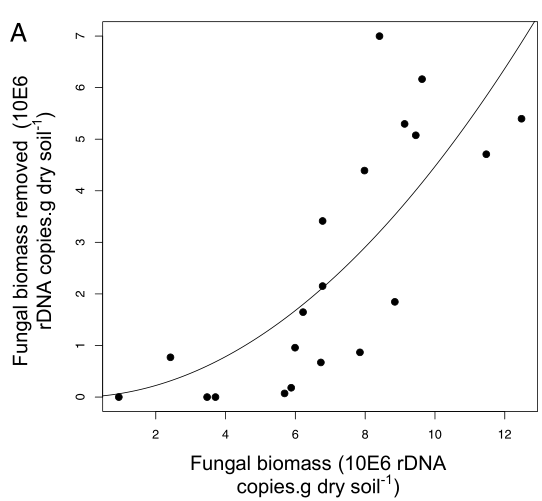
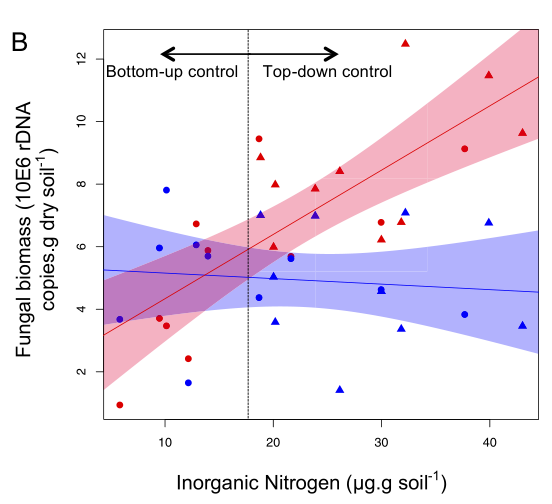
Fig. 4. Functional response of isopods along a gradient of fungal resource availability (A) and the consequences for fungal growth along a gradient of soil inorganic nitrogen availability (B). A shows how isopod grazing damage (estimated by subtracting fungal biomass in +fungal cords,−isopods chambers from that in the +fungal cords,+isopods plots), varies along a continuous gradient of resource availability. The initial lag in consumption rates is indicative of a type III functional response, as proportional grazing rates increase beyond a certain threshold of fungal availability. B shows changes in fungal biomass along an inorganic nitrogen gradient in the absence (red) and presence (blue) of grazing isopods. There were negligible effects of isopods on fungal biomass at ambient levels of nitrogen availability, but increasing fungal availability stimulated the proportional effects of grazing, mitigating the effect of nitrogen enrichment. Dots and triangles represent fungal biomass values in ambient and nitrogen-addition plots, respectively.
There were no effects of grazing below a fungal biomass threshold of ∼6 μg/g soil, but the proportion of fungus removed by grazers increased with fungal biomass. The top-down control emerges as a regulatory force across a gradient of soil nitrogen availability. Under ambient values of inorganic nitrogen availability (0–20 μg/g soil), fungal growth was limited by soil nutrient availability and remained below the threshold for isopod grazing effects. However, as increasing inorganic nitrogen availability alleviated the nutrient limitation on fungal cord growth, increasing rates of grazing damage allowed grazing to emerge as the dominant control on decomposer fungal activity.
Conclusions:
We show that trophic interactions in soil can regulate ecosystem-level responses to global change, even within a highly complex natural food web. Contemporary theory posits that top-down control of soil communities is apparent only when microbial activity is not constrained by other limiting factors (20, 28, 29). We show that, when global change factors alleviate the bottom-up constraints on microbial activity, top-down control is likely to emerge as an increasingly important ecological force. The stabilizing effects of such biotic interactions may contribute to the eventual subsidence of carbon cycling rates observed following initial increases in microbial activity in long-term climate change experiments (10, 15). Global change studies that alleviate the temperature, moisture, or nutrient limitations on microbial activity without incorporating top-down interactions are likely to inflate the projected strengths of carbon cycle–climate feedbacks.
Chen Yunru