原名:Stream microbial communities and ecosystem functioning show complex responses to multiple stressors in wastewater
译名:河流微生物群落和生态系统功能表现出对废水中多种应激源的复杂反应
期刊:Global Change Biology (Top1区,IF:8.55)
发表时间:2020年10月
关键词:生物多样性,碳处理,棉条实验,微量污染物(MPs),下一代测序,营养,温度,变暖
DOI:https://doi.org/10.1111/gcb.15302
Abstract Multiple anthropogenic drivers are changing ecosystems globally, with a disproportionate and intensifying impact on freshwater habitats. A major impact of urbanization are inputs from wastewater treatment plants (WWTPs). Initially designed to reduce eutrophication and improve water quality, WWTPs increasingly release a multitude of micropollutants (MPs; i.e., synthetic chemicals) and microbes (including antibiotic-resistant bacteria) to receiving environments. This pollution may have pervasive impacts on biodiversity and ecosystem services. Viewed through multiple lenses of macroecological and ecotoxicological theory, we combined field, flume, and laboratory experiments to determine the effects of wastewater (WW) on microbial communities and organic-matter processing using a standardized decomposition assay. First, we conducted a mensurative experiment sampling 60 locations above and below WWTP discharges in 20 Swiss streams. Microbial respiration and decomposition rates were positively influenced by WW inputs via warming and nutrient enrichment, but with a notable exception: WW decreased the activation energy of decomposition, indicating a “slowing” of this fundamental ecosystem process in response to temperature. Second, next-generation sequencing indicated that microbial community structure below WWTPs was altered, with significant compositional turnover, reduced richness, and evidence of negative MP influences. Third, a series of flume experiments confirmed that although diluted WW generally has positive influences on microbial-mediated processes, the negative effects of MPs are “masked” by nutrient enrichment. Finally, transplant experiments suggested that WW-borne microbes enhance decomposition rates. Taken together, our results affirm the multiple stressor paradigm by showing that different aspects of WW (warming, nutrients, microbes, and MPs) jointly influence ecosystem functioning in complex ways. Increased respiration rates below WWTPs potentially generate ecosystem “disservices” via greater carbon evasion from streams and rivers. However, toxic MP effects may fundamentally alter ecological scaling relationships, indicating the need for a rapprochement between ecotoxicological and macroecological perspectives.
KEYWORDS biodiversity, carbon processing, cotton-strip assay, micropollutants, next-generation sequencing, nutrients, temperature, warming
Main conetnts:
Cotton-strip assay:
Twenty study sites located across Switzerland were selected to investigate WWTP impacts on receiving stream ecosystems (Burdon et al. 2016, Stamm et al. 2016). At each study site, we designated one downstream sampling location (D), and two upstream sampling locations (U1, U2; Stamm et al. 2016). Location D was selected so that discharged WW was completely mixed across the wetted channel. Location U1 was chosen as close to the discharge as possible, and U2 approximately equidistant to that between U1 and D.
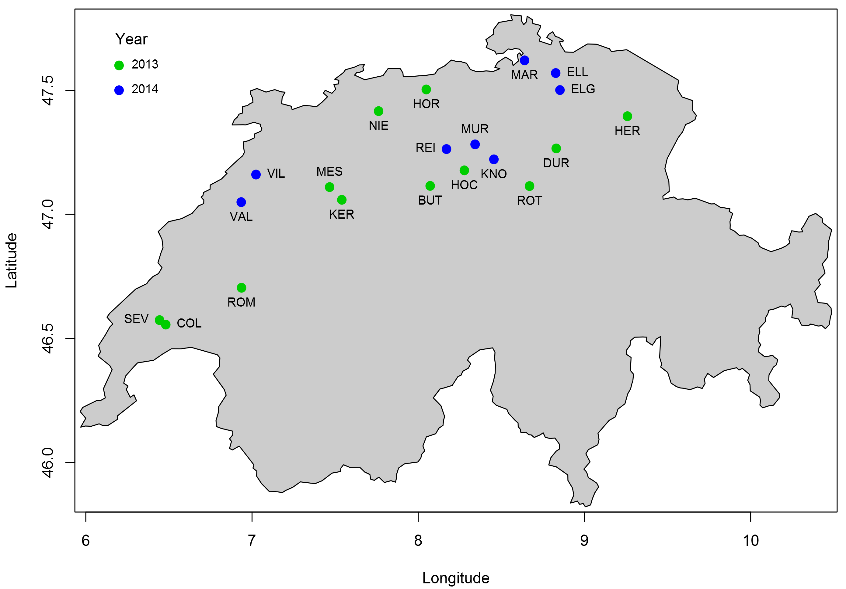
Map of study sites in Switzerland where the cotton-strip assay was deployed during the field experiment (12 sites in 2013, and 8 sites in 2014).
Flumes experiments:

Results:
Wastewater impacts on microbial communities
Bacterial diversity and composition on CSs responded to WW inputs. Rarefied bacterial OTU richness decreased at downstream sampling locations (Table 2), but the difference was not statistically significant. Instead, we saw a negative influence of non-fungicides TUs countered by a positive influence of nutrients (Table SD7). Likewise, moderate effect sizes indicated that WW presence had a positive influence on dominant taxa (Berger–Parker index) and a negative influence on rare taxa (Fisher's alpha), but these indicators along with Shannon diversity and Pielou's evenness showed no statistically significant changes (Table 2). Our analysis of bacterial OTU composition showed that downstream communities were significantly different based on presence–absence (“adonis,” F1,17 = 1.11, p < .05, R2 = 0.058; Figure 1a) and relative-abundance data (F1,17 = 0.83, p < .05, R2 = 0.045; Figure SD1). However, only NMDS2 based on relative-abundance data showed a significant difference between upstream and downstream sampling locations bacterial taxa responding negatively to WW included Lacihabitans (Figure 1a), and taxa in the Rhodocyclales and Burkholderiales (Table SD5). Beta-diversity partitioning indicated that a far greater proportion of community dissimilarity at downstream sites was attributable to turnover of OTUs (Simpson’s index 0.205 ± 0.049, 1 SD), as compared with the pure loss (or gain) of OTUs (Nestedness index 0.072 ± 0.065).
Fungal diversity and composition on CSs responded to inputs of WW. The presence of WW had a significantly negative effect on fungal α-diversity, as indicated by a decrease in OTUs at downstream locations (Table 2), and there was evidence for negative influences of fungicide TUs (Table SD7). Using presence–absence data, we found that there was a shift in fungal community composition at the downstream locations (“adonis,” F1,17 = 0.981, p < .05, R2 = .052; Figure 1b). Our NMDS analyses (presence–absence data) revealed that only NMDS2 showed a significant difference between upstream and downstream (Table 2). Redundancy analysis indicated that MPs and temperature jointly influenced fungal community composition (Table SD9). An unidentified species in the Trichosporonaceae was a potential fungal indicator taxon responding positively to WW inputs, whereas Vishniacozyma responded negatively (Figure 1b; Table SD6). Despite the loss of α-diversity, β-diversity partitioning within sites indicated that a greater proportion of community dissimilarity was attributable to turnover of OTUs (Simpson’s index 0.376 ± 0.098, 1 SD) compared with the pure loss (or gain) of OTUs (Nestedness index 0.054 ± 0.087).
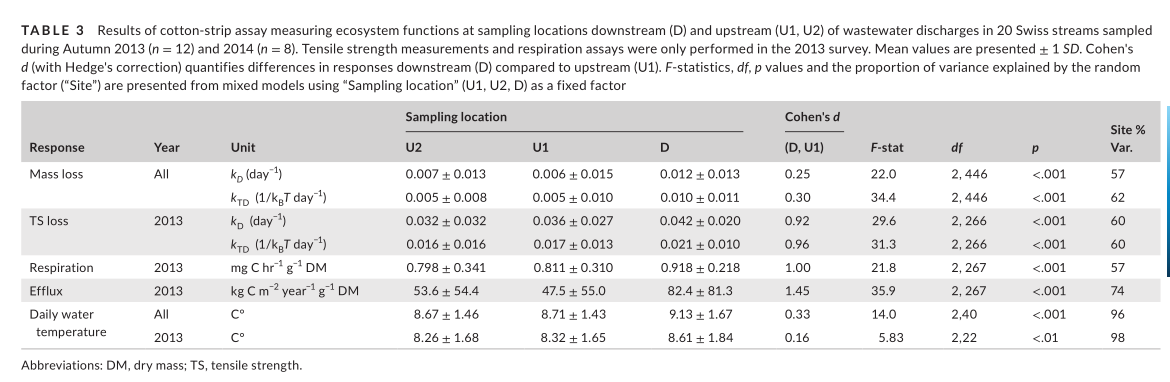
Overall, we found that within-site changes to fungal community composition were greater than bacteria with significantly higher turnover in response to WW (t19 = −10.8, p < .001). Despite this result, cotton decomposition rates (TS loss) were more strongly associated with bacterial (“rda,” F1,18 = 3.65, p < .05, R2 adj = .12) than fungal (F1,18 = 1.84, p < .05, R2 adj = .04) community composition. Finally, although we were unable to detect significant differences in community dispersion between sampling locations, we found evidence for sampling location-dependent changes in β-diversity across sites using beta-diversity partitioning. Fungal and bacterial communities at WW-impacted locations showed greater nestedness and overall dissimilarity (“betapart,” p < .001) across sites than upstream communities, indicating the potential for WW to contribute to changes in regional diversity patterns (γ-diversity).
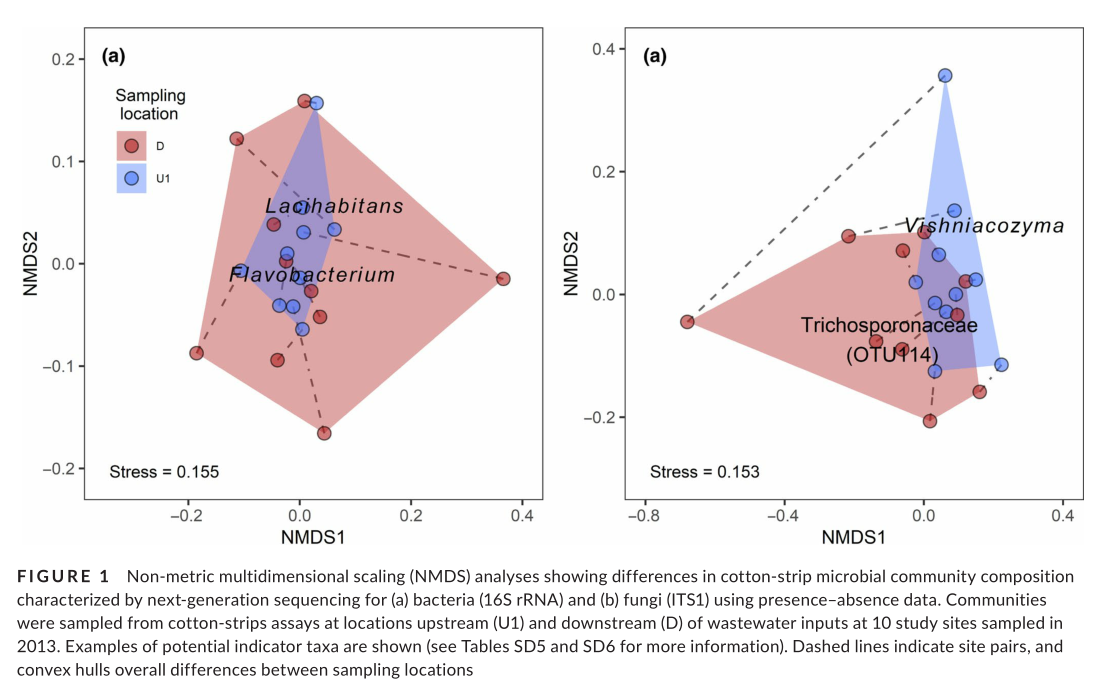
Experimental evidence of altered functioning
In the flumes experiment with fractions of treated WW (Exp. 1), we found that diluted WW inputs generally increased respiration and TS loss rates (Figure 4a,b). Respiration was the most sensitive response (F3,53 = 118, p < .001), indicating significant differences between all treatments except between the low and medium WW fractions (Figure 4a). Respiration rates at the highest fraction of WW (85%) were lower than that observed in the 15% and 50% WW treatments. In contrast, cotton breakdown responded positively in all WW fractions as demonstrated by TS loss rates (F3,56 = 89.5, p < .01), but there were no significant differences between the medium WW fraction (50%) and the other WW dilutions (Figure 4b). Similar results were observed for mass loss rates (X2 3 = 120, p < .001).
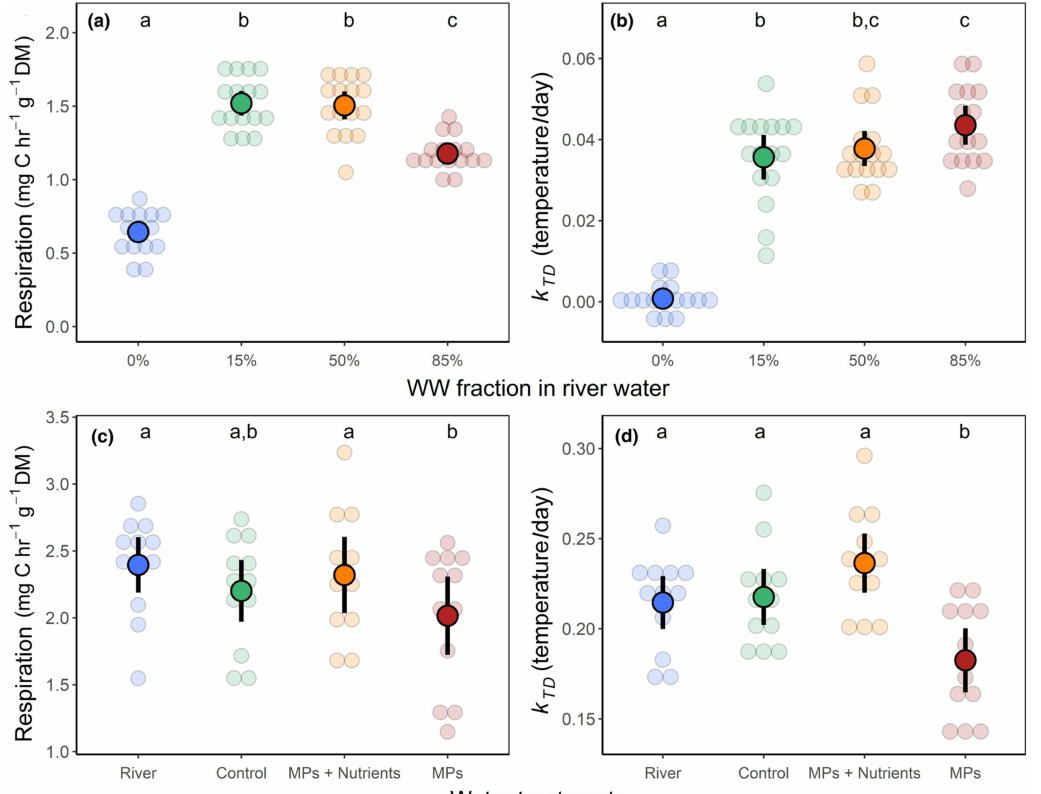
Exp. 2 results are reported in Appendix D because treatments were affected by the methanol carrier. In Exp. 3, we found that MPs had a significantly negative effect on ecosystem functioning as measured by microbial respiration (F3,43 = 8.06, p < .001; Figure 4c), cotton TS loss (F3,41 = 8.36, p < .001; Figure 4d), and mass loss (F3,40 = 11 . 2 , p < . 0 01). There were no significant differences between the “River,” “Technical Control” (i.e., the MeOH carrier treatment), and the combined “MPs and Nutrients” treatment (Figure 4c,d). Respiration rates in the technical control and MPs treatments were not significantly different (“lsmeans,” t = 2.37 , p = .088). However, we did find that comparing effect sizes relative to the river treatment indicated a stronger negative effect in the “MPs” treatment (Cohen's D ± 1 SE, −0.97 ± 0.50) than in the technical control (−0.49 ± 0.41). Furthermore, we found a small negative effect of the MPs treatments relative to the technical control (−0.33 ± 0.35). Results in Exp. 3 were similar to those in Exp. 2 (Appendix D.2).
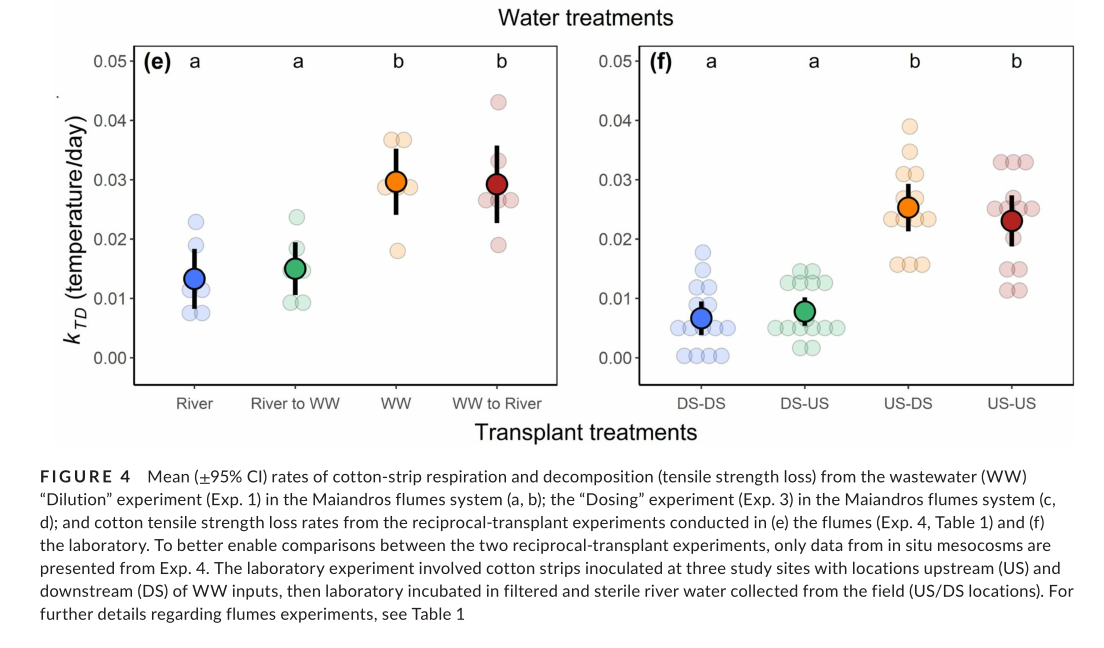
The flumes transplant experiment (Exp. 4) showed that inoculation with WW-borne microbes led to greater cotton TS loss (Figure 4e) that persisted when strips were transferred to river water (t = 0.14, p = .89). Likewise, the strips inoculated in river water showed no significant change when transferred to diluted WW (t = −1.13, p = .54). We observed virtually identical responses with mass loss, except for a small relative decrease (−19%) when strips inoculated in WW were transferred to river water (t = 3.88, p < .001), and a larger relative increase (38%) when strips inoculated in river water were transferred to diluted WW (t = −5.23, p < .001). These responses may have been due to differences in nutrient concentrations, although caution is needed when interpreting mass-loss data (Colas et al., 2019).
Concluding remarks
We used a wide range of approaches to study the effects of WW and associated stressors on a fundamental stream ecosystem process (i.e., organic-matter decomposition) using the CSA. Our study shows that WW-born MPs have the potential to harm ecosystem processes as indicated by the CSA, but these impacts may be “masked” by nutrients, warming, and stress-tolerant microbes. While we did find significant net positive effects of WW inputs on this functional indicator, the “slowing” of responses to warming across the temperature gradient indicates that the buffering capacity of microbial communities may be limited. This result, coupled with negative influences of fungicides on fungal richness, add to the growing concern about the threat this pesticide class poses to biodiversity and ecosystem functioning (Zubrod et al., 2019). Our findings point to the need for further experiments to manipulate abiotic and biotic drivers to better understand ecological limits and underlying mechanisms of change in freshwater ecosystems (Burdon et al., 2016).
Wastewater treatment plants are vitally important infrastructure for managing water resources (Schwarzenbach et al., 2010), and several developed countries are upgrading their facilities to help mitigate the growing threat from MPs (Stamm et al., 2016). Measuring ecological responses to upgraded infrastructure indicates the return of these investments and helps underscore the societal relevance of our research. Our results suggest that the subtle negative effects of MPs on microbial-mediated decomposition could be ameliorated with tertiary treatment steps (e.g., ozonation) that remove MPs. Thus, decomposition rates measured by the CSA should increase post-upgrade, signifying the potential for this functional indicator as a useful baseline to assess the efficacy of mitigation. However, such improvements may generate undesirable side effects. Our estimates suggest that WWTP inputs led to a median 39% increase in CO2 evasion rates. This efflux meant that in Switzerland there could be an additional 30,000 tons of carbon entering the atmosphere annually from WW-impacted streams and rivers, with the potential to further increase with improved treatment that removes MPs.
In a broader sense, our findings demonstrate the value of the CSA as a standardized method to measure microbial communities and organic-matter decomposition, helping to emphasize its utility as a functional indicator for ecosystem assessment (Colas et al., 2019; Tiegs et al., 2019). We found that the CSA was highly sensitive to a variety of environmental factors across field, flumes, and laboratory settings. Future research should describe microbial biodiversity in more detail to map structure to function, and further compare the relative performance of this functional indicator with natural leaf litter and other standardized organic-matter surrogates. Integrating knowledge of structural changes in stream communities with functional consequences will help to predict the impacts of synthetic chemicals and other global change drivers. Overall, our study further highlights the need to manage for multiple stressors when considering human impacts on the environment. Finally, we conclude that further reconciling macroecological and ecotoxicological perspectives is required to develop the transdisciplinary approach needed to understand interactions between environmental stressors, biodiversity change, and ecosystem functioning.
创新点:
CSA对温度、营养和污水输入高度敏感,最终结果显示污水处理厂下游的微生物活性增加,可能导致更大的CO2流出。
来源于污水的MPs(微量污染物)可能会损害生态系统过程,但这些影响可能被营养物质、变暖和耐压力的微生物“掩盖”。
CSA是衡量微生物群落和有机物质分解的可行方法。
Contact: Liu Run
E-mail: LiuRun24@outlook.com