A novel biosensor to monitor proline in pea root exudates
and nodules under osmotic stress and recovery
Title: A novel biosensor to monitor proline in pea root exudates and nodules under osmotic stress and recovery
Download website:
https://doi.org/10.1007/s11104-020-04577-2
Background and aims
Plant and bacteria are able to synthesise proline, which acts as a compound to counteract the negative effects of osmotic stresses. Most methodologies rely on the extraction of compounds using destructive methods. This work describes a new proline biosensor that allows the monitoring of proline levels in a non-invasive manner in root exudates and nodules of legume plants.
Methods
The proline biosensor was constructed by cloning the promoter region of pRL120553, a gene with high levels of induction in the presence of proline, in front of the lux cassette in Rhizobium leguminosarum bv. viciae.
Results
1. The proline biosensor OPS0650 is able to detect concentrations of proline in the nanomolar range
To determine the specificity of the induction of lux expression in the biosensor, bacterial cells were grown in UMS supplemented with different compounds and luminescence was measured after 3 h of incubation (Fig.1). Specific luminescence data are represented as relative luminescence units per optical density at 600 nm (RLU/OD600) to account for the differences in bacterial growth. The proline biosensor showed luminescence values 86-fold higher in the presence of 500 µM of proline than those to cells grown in control medium. Cells grown in media supplemented with 10 mM of L-4-hydroxyproline, a closely related non-proteinogenic amino acid, presented luminescence values comparable to those of a culture grown in the absence of proline, suggesting that the luminescence recorded is specific for proline. When pyruvate was eliminated from the media, the luminescence showed a decline, yet was still 17-fold higher than that of cells grown in the absence of the amino acid (Fig.1).
To define the limit of detection of the biosensor, the strain was incubated in UMS media supplemented with different proline concentrations ranging from 1 nM to 500 µM (Fig.1). The biosensor was able to detect concentrations of proline as low as 300 nM, showing a significant specific luminescence of around 4.42 × 105 ± 0.82 × 105 RLU/OD600. Increases in the concentration of proline over 50 µM did not produce significant increments in luminescence emission, maintaining luminescence values of approximately 5.63 × 106 ± 8.22 × 105 RLU/OD600.
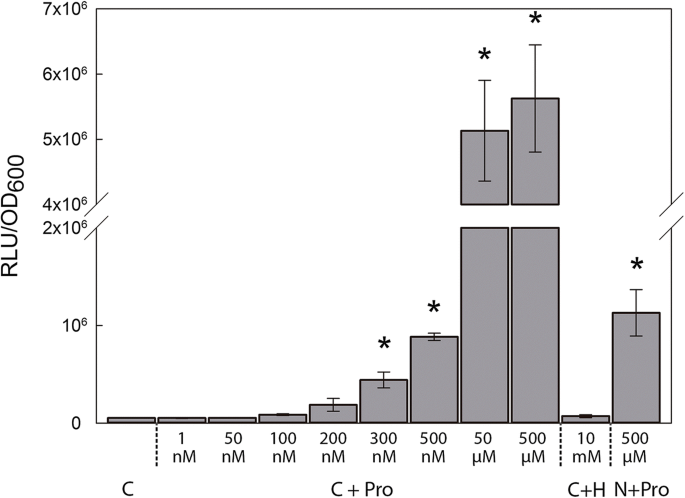
Fig.1 Specificity and sensitivity of the proline biosensor strain OPS0650. Luminescence values are expressed as relative luminescence units (RLU) per optical density at 600 nm (OD600). Cells were grown using different combinations of universal minimal salts (UMS) media. C stands for control growth media containing UMS + 30 mM pyruvate + 10 mM ammonium chloride (NH4Cl); C + Pro, control medium supplemented with various concentrations of proline as stated; C + H, control medium + 10 mM L-4-hydroxyproline; N + Pro, UMS medium + 10 mM NH4Cl + 500 µM Pro. Values represent mean ± SE from two independent experiments in the case of nM proline concentrations and three independent experiments in the other measurements. An asterisk (*) indicates significant differences from C (Student´s t-test at p ≤ .05)
2. In vivo monitorisation of proline in root exudates and nodules
We then analysed the spatial and temporal expression of the lux reporter in plants inoculated with the biosensor strain during root growth and nodule development. Pea plants were inoculated with either the biosensor strain OPS0650 or the negative and positive control strains LMB542 and D5250, respectively. Luminescence was measured at different time points from 4 to 20 dpi (Fig.2). Four days after inoculation, luminescence was induced mainly on primary roots, particularly in the upper half section of the main root (Fig.2a). As lateral roots developed, luminescence could also be observed in the oldest lateral roots, although at relatively lower levels (Fig.2b-f). These discrepancies may be related to the inoculation procedure, which was carried out when plants presented only a main root. At 10 dpi luminescence was detected in nodules (Fig.2c), reaching maximum intensity values at 15 dpi (Fig.2d).
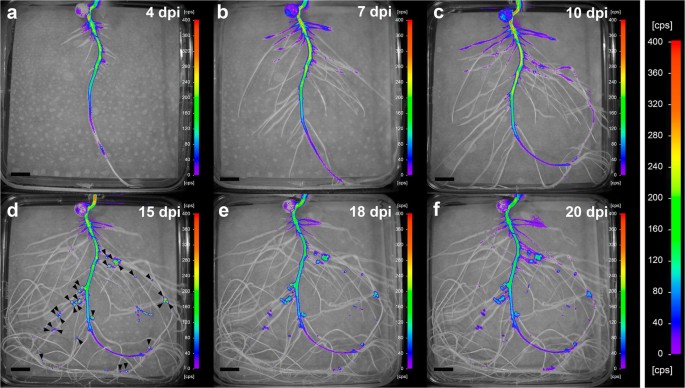
Fig.2 In vivo temporal and spatial expression of the proline biosensor strain in pea roots and nodules. Images are representative of plates corresponding to 5 biological replicates and were acquired at 4 (a), 7 (b), 10 (c), 15 (d), 18 (e), and 20 (f) dpi. Nodules were visible to the naked eye at around 10 dpi. Arrowheads were added in image (d) to show nodule position. Note that the same scale has been used in all images to facilitate comparisons. Scale bar, 1 cm
3. Induction of the proline biosensor occurs after a water-deficit and osmotic stress period
To monitor the level of the water-deficit stress imposed, we checked the following physiological parameters: leaf water potential (Ψleaf), stomatal conductance, net photosynthesis and symbiotic nitrogen fixation (Tab.1). At this stage (16 dpi), Ψleaf showed a significant decline, with values around − 0.49 ± 0.1 MPa, while control plants maintained at optimal water conditions showed Ψleaf values of -0.24 ± 0.05 MPa. Similarly, water deficit caused a 66% decline in stomatal conductance and a 36% reduction in photosynthetic rates compared to control plants. Regarding nitrogen fixation, water deficit and salt stress caused a 52 and 67% reduction in the rates of nitrogen fixation, respectively, compared to plants under optimal conditions (Tab.1).
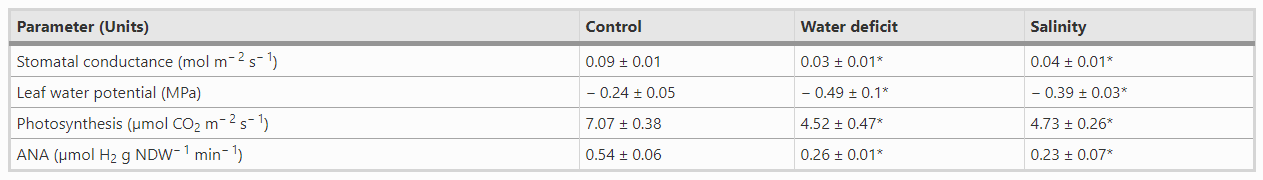
Tab.1 Effect of water deficit and osmotic stress on leaf water potential, stomatal conductance, photosynthesis and ANA of pea plants. Values represent the mean ± SE (7 ≤ n ≤ 10 biological replicates, except in ANA with n = 4). An asterisk (*) indicates significant differences compared to control plants. (Student´s t-test at p ≤ .05). NDW, nodule dry weight
Regarding the effect of the stresses on nodules, during the period of gradual water loss, the luminescence of nodules was maintained at relatively constant values, suggesting that proline levels did not show significant changes in bacteroids (Fig.3a, Fig.S4). At 16 dpi, plants were transferred to fresh plates (i.e., optimal water availability) as a recovery treatment. Recovery provoked a rapid increase in the luminescence observed in nodules within the first 24 h, showing a gradual reduction in the following days (Fig.3a and c, Fig.S1), almost reaching the levels of lux expression in nodules of control plants (Fig.3a and b).
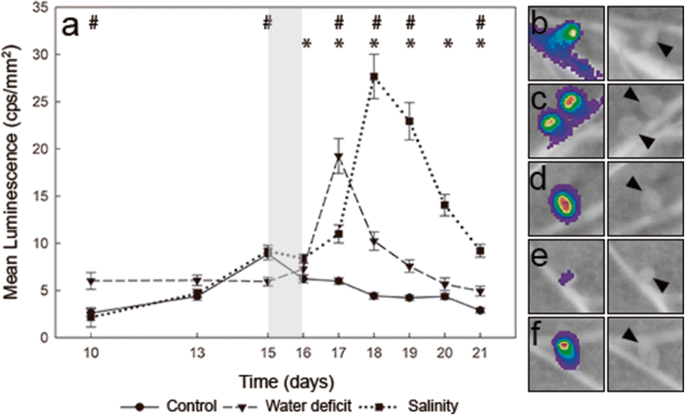
Fig.3 In vivo monitoring of proline in pea nodules of plants inoculated with the biosensor R. leguminosarum bv. viciae strain OPS650. Values represent mean luminescence [counts per second (cps) mm− 2] from nodules of pea plants inoculated with the proline biosensor. Stress was induced at 15 dpi for 24 h (grey area). For recovery, plants were transferred to fresh plates (optimal growth conditions). Values represent the mean ± SE calculated from 5 biological replicates using the luminescence values of all the nodules present in a plant. An asterisk or a hash sign (* or #) indicate significant differences between salt-stressed or water-deficit plants and control plants, respectively (ANOVA at p < .05, Dunnett T3 post hoc test). (b) to (f) representative images from nodules of pea plants under different conditions, and their corresponding luminescence image. b nodules of plants inoculated with the proline biosensor strain OPS0650 under control conditions (17 dpi, scale 0-400 cps); c nodules from water-deficit plants at the maximum level of luminescence of the proline biosensor during recovery (17 dpi, scale 0-1100 cps); d nodules from salt-stressed plants at the maximum luminescence of the proline biosensor during recovery (18 dpi, scale 0-3000 cps); e nodules from plants inoculated with the negative control strain LMB542 (no lux expression, 17 dpi, scale 0-400 cps); f nodules from plants inoculated with the positive control strain D5250 (constitutive lux expression,17 dpi, scale 1200–25200 cps). Arrowheads were added in images (b) to (f) to show nodule position
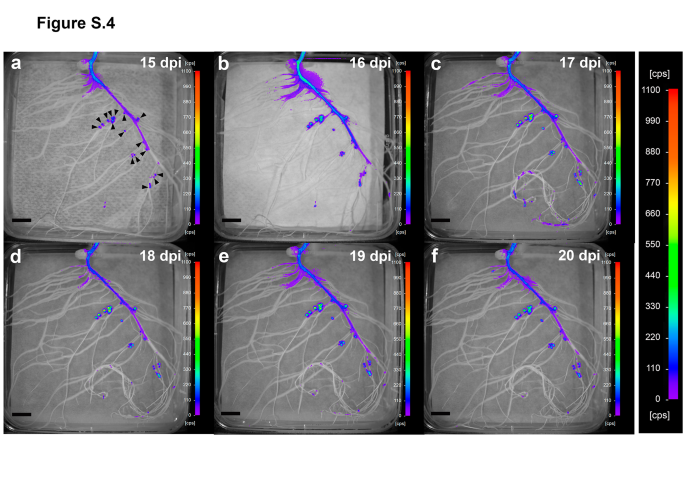
Fig. S4 In vivo temporal and spatial expression of the proline biosensor strain in roots and nodules of water-stressed pea plants. Luminescence is expressed in counts per second (cps). Images were acquired at 15 (a), 16 (b), 17 (c), 18 (d), 19 (e), and 20 (f) dpi. Arrowheads were added in image (a) to show nodule position. Note that the same scale has been used in all images to facilitate comparisons. Representative images of plates belonging to a time series experiment (n = 5 biological replicates). Scale bar, 1 cm (PNG 6376 kb)
Salt stress was generated by incubating plants on plates containing 150 mM NaCl for one day. Similarly, to the water-deficit experiment, recovery was carried out by transferring the plants to fresh plates under optimal growth conditions, corresponding to the time point 16 dpi. Salinity also had a negative impact on the physiology of the plants, which showed Ψleaf values of -0.39 ± 0.03 MPa, a 55% decline in stomatal conductance and a 35% reduction in net photosynthesis (Tab.1). 24 h after the application of the salt stress there were no significant changes in the level of luminescence of the proline biosensor compared to the luminescence recorded before the onset of the treatment (Fig.3a, Fig.S5). At 17 dpi, however, luminescence started to increase, reaching its maximum at 18 dpi (Fig.3a and d, Fig.S5). Subsequently, the levels of luminescence progressively declined, yet were significantly higher than those of control nodules at the end of the experiment (Fig.3a).
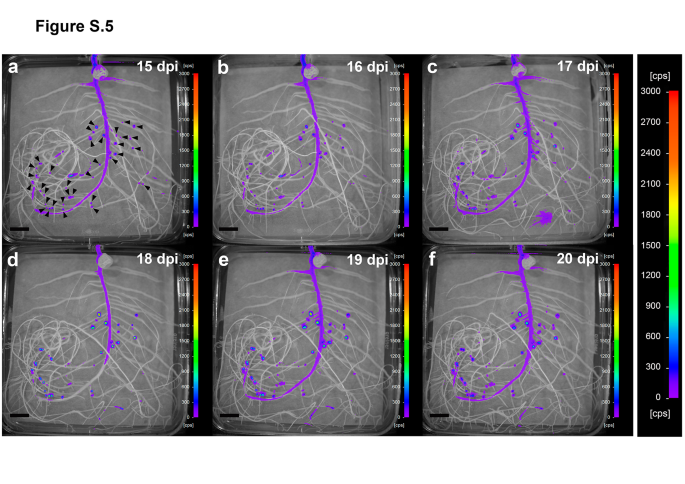
Fig. S5 In vivo temporal and spatial expression of the proline biosensor strain in roots and nodules of salt-stressed pea plants. Luminescence is expressed in counts per second (cps). Images were acquired at 15 (a), 16 (b), 17 (c), 18 (d), 19 (e), and 20 (f) dpi. Arrowheads were added in image (a) to show nodule position. Note that the same scale has been used in all images to facilitate comparisons. Representative images of plates belonging to a time series experiment (n = 5 biological replicates). Scale bar, 1 cm (PNG 6376 kb)
Conclusions
This biosensor is a useful tool for the in vivo monitoring of proline levels in root exudates and bacteroids of symbiotic root nodules, and it contributes to our understanding of the metabolic exchange occurring in nodules under abiotic stress conditions.
Contact: Liang Jiawen
E-mail: jwliang@smail.nju.edu.cn