Climate and litter traits predominantly control litter decomposition
Article 1:
Title: Traits drive global wood decomposition rates more than climate
https://onlinelibrary.wiley.com/doi/10.1111/gcb.14357
Abstract:
Wood decomposition is a major component of the global carbon cycle. Decomposition rates vary across climate gradients, which is thought to reflect the effects of temperature and moisture on the metabolic kinetics of decomposers. However, decomposition rates also vary with wood traits, which may reflect the influence of stoichiometry on decomposer metabolism as well as geometry relating the surface areas that decomposers colonize with the volumes they consume. In this paper, we combined metabolic and geometric scaling theories to formalize hypotheses regarding the drivers of wood decomposition rates, and assessed these hypotheses using a global compilation of data on climate, wood traits, and wood decomposition rates.Our results are consistent with predictions from both metabolic and geometric scaling theories. Approximately half of the global variation in decomposition rates was explained by wood traits (nitrogen content and diameter), whereas only a fifth was explained by climate variables (air temperature, precipitation, and relative humidity). These results indicate that global variation in wood decomposition rates is best explained by stoichiometric and geometric wood traits. Our findings suggest that inclusion of wood traits in global carbon cycle models can improve predictions of carbon fluxes from wood decomposition.
Main content:

FIGURE 1 Variation in wood decomposition rates at 191 sites spanning global climate gradients. (a) Geographical distribution of the sites where data were available in forest distribution map (Crowther et al., 2015). (b) Variation in annualized wood decomposition rates (k, year−1) in relation to mean annual temperature and mean annual precipitation. All major terrestrial plant biomes are shown (Whittaker, 1970). Circle size and color represent classes of decomposition rates. (c) Correlations between annualized wood decomposition rates (k, year−1) and variables of climate and wood traits by multiple regression. Positive correlations are displayed in blue and negative correlations in red color. Color intensity and the size of the circle are proportional to the correlation coefficients. Variable names: k, wood decomposition rate (year−1); N, wood N content (g N/g M); D, wood diameter (cm); ρ, wood mass density (g/cm3); <1/kBT >ma, averaged annual temperature (eV−1); Pma, mean annual precipitation (mm); hma, mean annual relative humidity (%); las, length of the active season (d); <1/kBT >as, averaged active season temperature (eV−1); Pas, active season precipitation (mm); and has, active season relative humidity (%).
TABLE 1 Coefficients, 95% confidence interval (95% CI), and r2values for ordinary least squares regression models, multiple regression models, and linear mixed‐effects models for global wood decomposition rates.
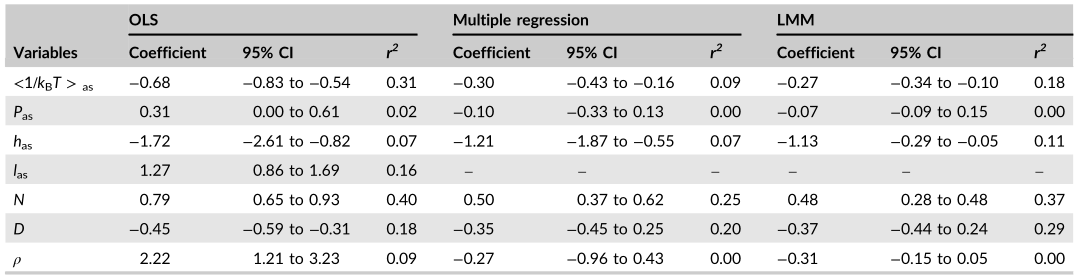
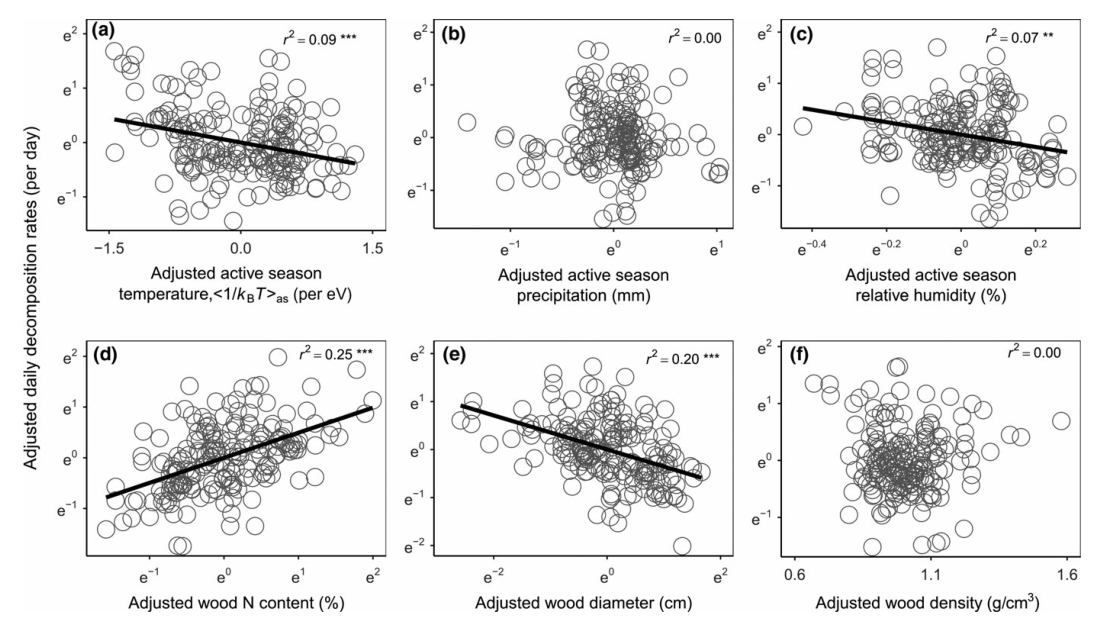
FIGURE 2 Partial regression plots illustrating relationships between active season wood decomposition rates and individual covariates from Equation (2) at 191 globally distributed sites. Plots show the relation (slope and variance) between decomposition rates and each covariate while controlling for the influence of all other model covariates. (a) Mean active season temperature; (b) Mean active season precipitation; c, Mean active season relative humidity; (d) Wood N content; (e) Wood diameter; (f) Wood density; (e) mathematical constant (~2.718). ** and *** indicate significant correlation between wood decomposition rates and the corresponding variable at p < 0.01 and p < 0.001, respectively. There were no significant linear relationships between active season decomposition rate and active season precipitation, and wood density.
FIGURE 3 Relative importance of climate and wood traits on controlling global wood decomposition rates (n = 191). (a) Model-averaged effect size of the predictors on active season wood decomposition rates (based on Z-scores with linear mixed-effects models) at the global scale. (b) Percentage of variations in active season decomposition rates explained by climate, wood traits, and both climate and wood traits at the global scale. Points denote average estimates, and lines denote 95% confidence intervals. Filled circles indicate significance at p < 0.05. Climate variables in skyblue; wood traits in green.
Conclusion:
In conclusion, our study combined MST and GST to link multiple hypothesized drivers to global variations in wood decomposition rates. Using multiple approaches to fit our theory to data spanning global climate gradients, we have shown that global variation in wood decomposition was consistent with predictions from both MST and GST. Our compiled data conformed to power laws as predicted by both theories. Additionally, multiple regression and mixed model estimates of the activation energies for wood decomposition were statistically indistinguishable from expectations for the kinetics of extracellular enzymes that are important for microbial decomposition. Furthermore, diameter‐scaling exponents estimated using three regression approaches were all consistent with how the scaling of length with diameter in decomposing wood particles departs from idealized optimality assumptions. We have also shown that wood traits were stronger predictors of variation in wood decomposition rates than climate variables. Diameter and wood N content were the most important traits controlling global normalized decomposition rates. Specifically, wood diameter influenced the relationship between the surface area on which microbial decomposers reside to the volume of the wood that they consume, resulting in a decrease in decomposition rates with wood diameter (Cornwell et al., 2009; van der Wal et al., 2007; van Geffen et al., 2010). The importance of wood N potentially originates from the relatively low nutrient availability of most dead wood. The observed increase in wood decomposition rates with N content has important implications for assessing effects of N deposition on wood decomposition rates. Although wood traits are key drivers of variation in decomposition rates, they are not included in most Earth system models, and most models have only one single coarse wood debris pool for wood decomposition (e.g., the Community Land Model; Oleson et al., 2013). Our results suggest that inclusion of wood traits in these models will improve global change predictions.
Article 2:
Title: Leaf litter traits predominantly control litter decomposition in streams worldwide
https://onlinelibrary.wiley.com/doi/full/10.1111/geb.12966
Abstract:
Aim: Leaf litter decomposition in freshwater ecosystems is a vital process linking ecosystem nutrient cycling, energy transfer and trophic interactions. In comparison to terrestrial ecosystems, in which researchers find that litter traits predominantly regulate litter decomposition worldwide, the dominant factors controlling its decomposition in aquatic ecosystems are still debated, with global patterns not well documented. Here, we aimed to explore general patterns and key drivers (e.g., litter traits, climate and water characteristics) of leaf litter decomposition in streams worldwide.
Location: Global.
Time period: 1977–2018.
Major taxa studied: Leaf litter.
Methods: We synthesized 1,707 records of litter decomposition in streams from 275 studies. We explored variations in decomposition rates among climate zones and tree functional types and between mesh size groups. Regressions were performed to identify the factors that played dominant roles in litter decomposition globally.
Results: Litter decomposition rates did not differ among tropical, temperate and cold climate zones. Decomposition rates of litter from evergreen conifer trees were much lower than those of deciduous and evergreen broadleaf trees, attributed to the low quality of litter from evergreen conifers. No significant differences were found between decomposition rates of litter from deciduous and evergreen broadleaf trees. Additionally, litter decomposition rates were much higher in coarse than in fine-mesh bags, which controled the entrance of decomposers of different body sizes. Multiple regressions showed that litter traits (including lignin, C:N ratio) and elevation were the most important factors in regulating leaf litter decomposition.
Main conclusions: Litter traits predominantly control leaf litter decomposition in streams worldwide. Although further analyses are necessary to explore whether commonalities of the predominant role of litter traits in decomposition exist in both aquatic and terrestrial ecosystems, our findings could contribute to the use of trait-based approaches in modelling the decomposition of litter in streams globally and exploring mechanisms of land–water–atmosphere carbon fluxes.
Main content:

FIGURE 1 The global distribution of leaf litter decomposition records in streams/rivers (n =1,591). There were 1,707 cases in total, with 116 cases having no latitude reported.
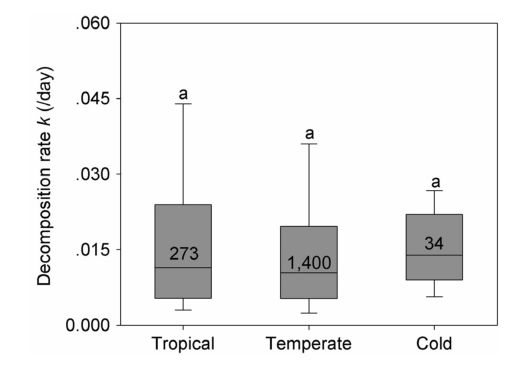
FIGURE 2 Comparison of leaf litter decomposition rate in streams (k, per day) among different climate zones. Sample size is indicated by the number inside of each column. Different lowercase letters on error bars indicate significant differences at p < .05

FIGURE 3 Comparison of leaf litter decomposition rate in streams (k, per day) among different tree functional types. DB = deciduous broadleaf; EB = evergreen broadleaf; EC = evergreen onifer. Sample size is indicated by the number inside of each column. Different lowercase letters on error bars indicate significant differences at p < .05.

FIGURE 4 Comparison of the litter decomposition rate in streams (k, per day) between coarse‐ and fine‐mesh size litter bags. Sample size is indicated by the number inside of each column. Different lowercase letters on error bars indicate significant differences at p < .05.
TABLE 1 Results of linear models for responses of litter decomposition rate in streams (k, per day) to climate zone, tree functional type, mesh size group and their interactions.
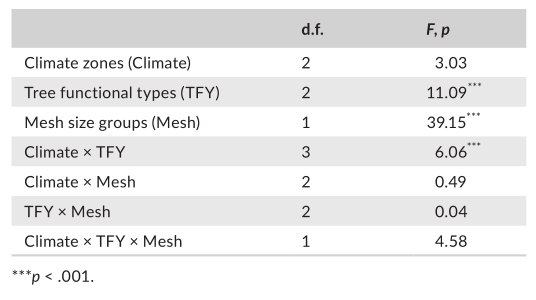
TABLE 2 Results of multi‐regression analyses of litter decomposition rate in streams (k, per day) with environmental factors, water characteristics of streams, litter traits and experimental factors.

Conclusion:
It is widely reported that litter decomposition is much faster in litter bags with a coarse rather than a fine‐mesh size (e.g., Gantes, Marano, & Rigacci, 2011; Ferreira, Chauvet, et al., 2015; Lecerf & Chauvet, 2008). This methodological aspect (mesh size) affects decomposition by means of interfering with decomposition processes, such as physical abrasion and decomposer activity, especially the feeding and maceration by leaf‐shredding invertebrates and microbial metabolism (Iñiguez‐Armijos et al., 2016; Langhans & T ockner, 2006; Stewart & Davies, 1989). Litter bags with coarse meshes allow large leaf‐consuming invertebrates to contact the litter, whereas those with fine meshes exclude a large portion of invertebrates without limiting microbial colonization (Lecerf & Chauvet, 2008). Fine mesh usually eliminates shredding by invertebrates and protects litter from heavy leaching and physical fragmentation, and litter decomposition is thus generally faster in the presence of macroinvertebrates than in their absence (e.g., Langhans & T ockner, 2006; Iñiguez‐Armijos et al., 2016; Roberts et al., 2016). Moreover, fungal growth and microbial colonization could be restricted by the protected environment (fine mesh), where exchanges of dissolved O2 and nutrients are reduced, which consequently retards the decomposition (Fleituch, 2001; LeRoy, Whitham, Keim, & Marks, 2006). A few studies have found no effect of mesh size on decomposition rates, and the authors attributed this to the extremely low litter quality and its associated macroinvertebrates, particularly shredders (Ágoston‐Szabó, Schöll, Kiss, & Dinka, 2016; Benfield, Paul, & Webster, 1979). Shredders prefer high‐quality litter, and their survivorship is low when fed with low‐quality litter (Canhoto & Graça, 1995). Together with the duration of decomposition, decomposers constitute significant factors influencing litter decomposition in streams. As the duration increases, for instance, this would contribute to the building of microbial assemblage composition, especially in the process of decomposition of conifer needles with thick cuticles (Newman, Liles, & Feminella, 2015). Many experiments choose the sampling time to match 50% litter mass loss, a time at which leaf litter reaches the peak ergosterol concentration (Ferreira, Chauvet, et al., 2015; Haapala et al., 2001). At the 50% breakdown point, leaf litter is most palatable to shredders, and shredder feeding is expected to be maximal (Cummins, Wilzbach, Gates, Perry, & T aliaferro, 1989; Richardson et al., 2004). Given the differences in the contents of nutrient elements and recalcitrant compounds of specific litter species, serious consideration should be given when choosing a suitable mesh size for litter bags and designing a rational sampling time.
In conclusion, our results showed that leaf litter traits predominantly controlled litter decomposition in streams worldwide, paralleling the findings for terrestrial ecosystems (Cornwell et al., 2008; Zhang et al., 2008). Our findings could contribute to the use of trait‐based approaches in modelling the decomposition of leaf litter in streams at the global scale and exploring mechanisms of land–water–atmosphere C fluxes. Further comprehensive analysis is required, however, to uncover whether commonalities of the predominant role of litter traits in decomposition exist in aquatic and terrestrial ecosystems, aimed at promoting the development of common global models.
Qi Chen