叶绿素a的变化途径——水生环境中浮游植物早期成岩过程中荧光叶绿素衍生物向非荧光叶绿素衍生物的逐步变化

1. 研究方法与背景
1.1 叶绿素a衍生物的层级划分
具有完整卟啉环结构的叶绿素主要包含3个官能团:—Mg、—COOCH3(甲酯基)、—phytyl(植醇)。叶绿素在降解过程中会逐渐脱去以上3个官能团:脱去Mg的叶绿素衍生物一般命名为脱镁——,脱去甲酯基则命名为为焦——,脱去植醇则命名为脱植醇——或——叶绿酸。在水体降解环境中,脱去以上3个基团的焦脱镁叶绿酸a往往为最后的绿色中间产物,再往后则分解为非荧光叶绿素衍生物(NCDs)。
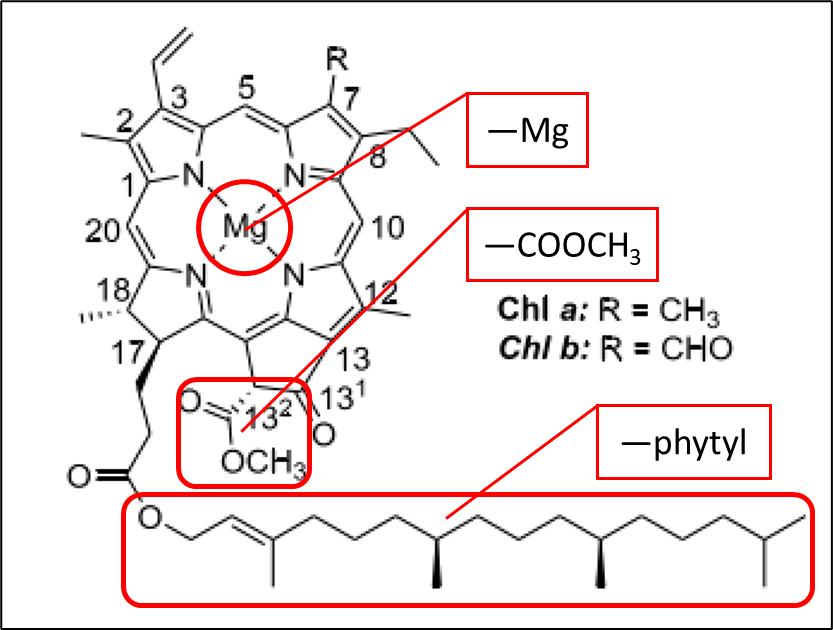
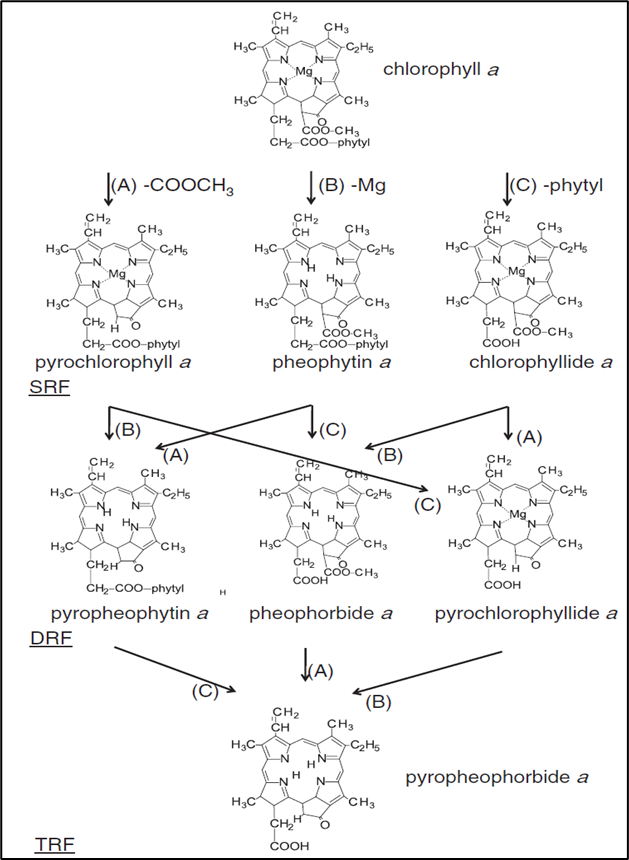
为了更好地对叶绿素a衍生物进行综合分析,根据叶绿素a衍生物中脱去官能团数量的不同,将叶绿素a衍生物分为SRF(单反应组分)、DRF(双反应组分)和TRF(焦脱镁叶绿酸a)三种存在形式。SRF指脱去1个官能团的叶绿素a衍生物,包括焦叶绿素a、脱镁叶绿素a和脱植基叶绿素a。DRF则代表脱去2个官能团的叶绿素a衍生物,包括焦脱镁叶绿素a、脱镁叶绿酸a和焦脱植基叶绿素a。TRF为脱去以上三个官能团的叶绿素a衍生物,一般为焦脱镁叶绿酸a,为最后的荧光性叶绿素衍生物,再往后则分解为非荧光性叶绿素衍生物(NCDs)。
1.2 叶绿素a荧光性衍生物和非荧光性衍生物的检测
高效液相色谱荧光检测法[1]可用于检测浮游植物降解后的叶绿素衍生物,但是该方法只能检测从叶绿素a到焦脱镁叶绿酸a在内的叶绿素衍生物,不能用于检测非荧光叶绿素衍生物(NCDs)。
将叶绿素衍生物经铬酸氧化后,其转化为EMMi(ethylmethylmaleimide,乙基甲基马来酰亚胺),EMMi的产量与叶绿素衍生物的吡咯环数量有关,不受荧光性的影响,因此通过检测EMMi的量可以反映出包括NCDs在内的叶绿素衍生物的总含量。本实验采用铬酸氧化 + 气相色谱-质谱联用仪(GC-MS)的方法[2]检测叶绿素衍生物的总含量。因此这两种方法的检测之差即为NCDs的含量。
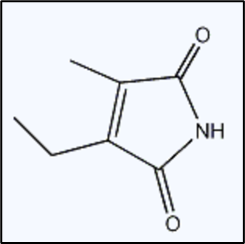
2-乙基-3-甲基马来酰亚胺
1.3 样品处理与材料准备
(1)样品处理:
包含天然微生物种群的表层海水于2010年7月8日和2011年4月18日从日本静冈市下田县乌拉湾(北纬34.7度,东经138.9度)获得,分别用于实验1和实验2。
样品经过富集后,将培养瓶放置在阳光下培养1.5天(记录为-1.5~0天),为光培养阶段。记录光照强度和温度等。而后转入黑暗中培养。
于第1、3、7、15、30、60和92天取样进行总叶绿素衍生物含量(铬酸氧化 + GC-MS)和荧光性叶绿素衍生物含量的测定(HPLC + 荧光检测)。
(2)材料准备:
叶绿素a、脱镁叶绿素a和脱植醇叶绿素a —— 购于Wako Pure Chemical Industries, Ltd.
脱镁叶绿酸a、焦脱镁叶绿酸a ——购于 TAMA Biochemical Co, Ltd.
焦叶绿素a、焦脱植醇叶绿素a、焦脱镁叶绿素a —— 自己制备[3](Pennington et al. 1964)
2. 主要结果
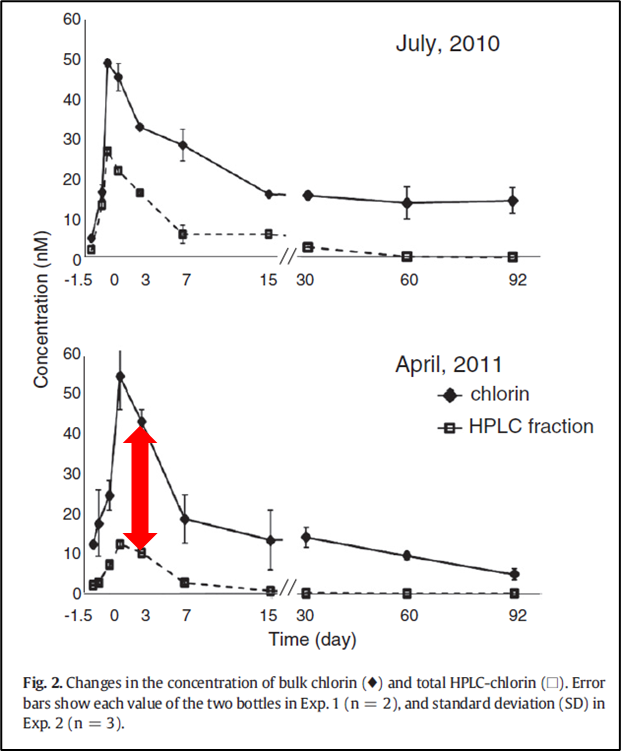
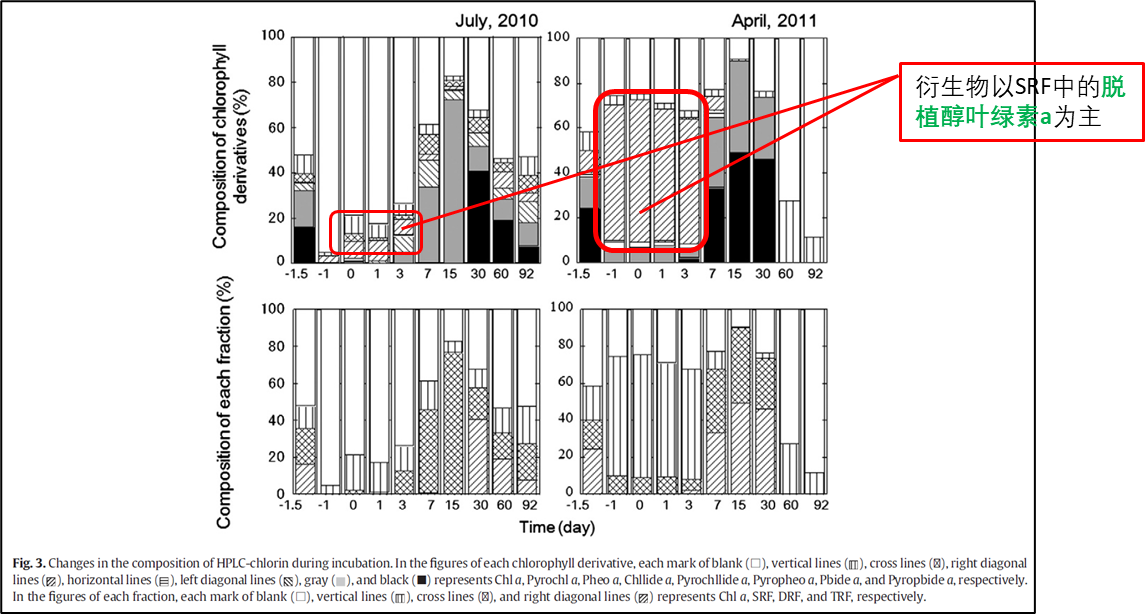
2.1 黑暗培养早期(第0—3天)
在黑暗培养的早期阶段,叶绿素a仍为主要成分,衍生物以SRF为主,其中以脱植醇叶绿素a为代表。有研究表明脱植基叶绿素a的产生可能与浮游植物自身叶绿素酶的活性有关[4][5][6],叶绿素酶的活性反映了浮游植物自身的生理条件,酶活性越高叶绿素a降解越早,因此其生理条件也就越差。
虽然两个实验都出现了SRF值的增加,但其贡献水平差异显著,实验2中的SRF比值显著高于实验1,这可能与浮游植物间的生理条件相关,表明实验1中浮游植物的生理条件优于实验2。
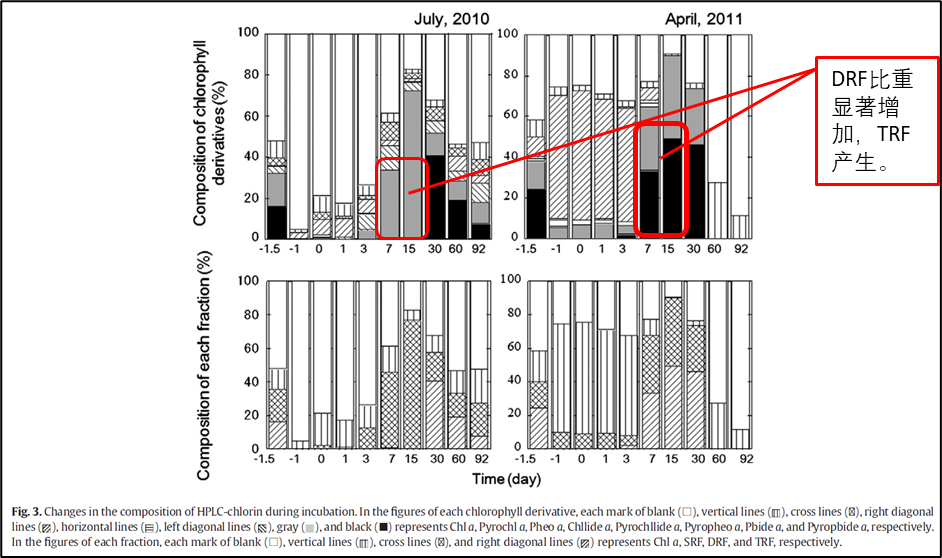
2.2 黑暗培养中期(第7—15天)
实验1中DRF的比重显著增加,实验2中DRF的比重显著增加的同时也出现了TRF。其中DRF中以脱镁叶绿酸a为主,TRF以焦脱镁叶绿酸a为主。有研究报道显示叶绿素a脱去两个基团需要有氧细菌的参与[7],因此DRF和TRF的积累与细菌的降解活动有关。
先前的研究表明,甲酯基在氧化条件下易于保存[8],但植醇侧链在缺氧条件下保存良好[9][10]。而脱镁叶绿素a由于甲酯基(—COOCH3)的存在,因此能在有氧条件下保存更久。而Mg、植醇侧链在有氧条件下易脱落。
2.3 黑暗培养晚期(第30—92天)
TRF在第15—30天显著增加,30天后,TRF的比重逐渐减少,在实验2中出现了明显的减少(第60天时)。TRF在实验2中的贡献比实验1中出现的更早且更显著,这可能与实验2中黑暗培养早期产生了更多的脱植基叶绿素a有关。
60天以后色素以叶绿素a为主,这可能与水体中矿物颗粒对残留叶绿素a的吸附作用有关,叶绿素长时间吸附在矿物表面时可以得到很好的保护[11][12]。虽然在这些实验所用的海水中可能只存在少量的矿物颗粒,但它可能足以保存少量的Chl a,为最大浓度的0.14%-0.6%。

2.4 叶绿素a的降解途径
浮游原生动物的放牧作用也被认为是影响海水中叶绿素降解的主要因素[13][14],原生动物放牧会在短期内产生大量脱镁叶绿酸a,而本实验脱镁叶绿酸a的出现时间较晚,因此可排除原生动物放牧作用的影响。
实验结果表明,叶绿素a的降解可以分为SRF、DRF和TRF三个组分阶段,这三个阶段的代表性主要产物分别为脱植基叶绿素a、脱镁叶绿酸a和焦脱镁叶绿酸a。因此综合以上结果可以得出水生环境中浮游植物早期成岩阶段叶绿素a的降解途径:即叶绿素a先脱去植醇侧链,再脱去Mg,最后脱去甲酯基。其中植醇侧链的脱去与浮游植物叶绿素酶的活性有关,其后的过程主要与细菌的降解作用有关。
2.5 NCDs的检测
本研究的结果显示,高效液相色谱-二氢卟吩浓度与铬酸氧化法测定的总衍生物浓度之间存在显著差异,这表明存在高效液相色谱无法表征的叶绿素衍生物。实验检测到了NCDs的产生,被认为是水柱早期成岩过程中的首例NCDs报告。在第15—90天持续检测到少量NCDs的存在,30天后总衍生物的含量没有显著变化,而荧光叶绿素衍生物逐渐消失,说明NCDs比其他叶绿素衍生物更难以被微生物降解,能够在水体中保存更长的时间。
3. 主要结论
(1)本研究报道了水体早期成岩过程中叶绿素a的降解途径,即脱植基——脱镁——脱甲酯基,降解为焦脱镁叶绿酸a,往后则降解为非荧光性叶绿素a衍生物。
(2)本研究采用铬酸氧化和GC-MS结合的方法将荧光性和非荧光性叶绿素衍生物的测定区分开来,对非荧光性叶绿素衍生物进行了定性的测定,表明其可能在水体中能保存较长时间不易被微生物降解。但是具体的降解途径没有进一步展开。
(3)叶绿素a在水体中的降解是浮游植物自身叶绿素酶和细菌降解活动综合作用下的过程。
参考文献
[1] Furuya, K., Hayashi, M., Yabushita, Y., 1998. HPLC determination of phytoplankton pigments using N, N-dimethylformamide. J. Oceanogr. 54, 199–203.
[2] Satoh, Y., Nomoto, S., Hama, T., 2012. Comprehensive determination of chlorophyll derivatives by chromic acid oxidation. Chem. Lett. 41, 571–573.
[3] Pennington, F.C., Strain, H.H., Katz, J.J., 1964. Preparation and properties of pyrochlorophyll a, methyl pyrochlorophyllide a, pyropheophytin a, and methyl pyropheophorbide a derived from chlorophyll by decarbomethoxylationl. J. Am. Chem. Soc. 86, 1418–1426.
[4] Jeffrey, S.W., Hallegraeff, G.M., 1987. Chlorophyllase distribution in ten classes of phytoplankton: a problem for chlorophyll analysis. Mar. Ecol. Prog. Ser. 35, 293–304.
[5] Louda, J.W., Li, J., Liu, L., Winfree, M.N., Baker, E.W., 1998. Chlorophyll a degradation during cellular senescence and death. Org. Geochem. 29, 1233–1251.
[6] Louda, J.W., Li, J., Liu, L., Baker, E.W., 2002. Senescence- and death-related alteration of chlorophylls and carotenoids in marine phytoplankton. Org. Geochem. 33, 1635–1653.
[7] Szymczak-Zyła, M., Kowalewska, G., Louda, J.W., 2008. The influence of microorganisms on chlorophyll a degradation in the marine environment. Limnol. Oceanogr. 53, 851–862.
[8] Keely, B.J., Brereton, B.G., Maxwell, J.R., 1988. Occurrence and significance of pyrochlorins in a lake sediment. Org. Geochem. 13, 801–805.
[9] Louda, J.W., Loitz, J.W., Rudnick, D.T., Baker, E.W., 2000. Early diagenetic alteration of chlorophyll a and bacteriochlorophyll a in a contemporaneous marl ecosystem; Florida Bay. Org. Geochem. 31, 1561–1580.
[10] Spooner, N., Getliff, J.M., Teece, M.A., Parkes, R.J., Leftley, J.W., Harris, P.G., Maxwell, J.R., 1995. Formation of mesopyrophaeophorbide a during anaerobic bacterial degradation of the marine prymnesiophyte Emiliania huxleyi. Org. Geochem. 25, 225–229.
[11] Hedges, J.I., Keil, R.G., 1995. Sedimentary organic matter preservation: an assessment and speculative synthesis. Mar. Chem. 49, 81–115.
[12] Keil, R.G., Montlucon, D.B., Prahl, F.G., Hedges, J.I., 1994. Sorptive preservation of labile organic matter in marine sediments. Nature. 370, 549–552.
[13] Shuman, F.R., Lorenzen, C.J., 1975. Quantitative degradation of chlorophyll by herbivore. Limnol. Oceanogr. 20, 580–586.
[14] Welschmeyer, N.A., Lorenzen, C.J., 1985. Chlorophyll budgets: zooplankton grazing and phytoplankton growth in a temperate fjord and the Central Pacific Gyres. Limnol. Oceanogr. 30, 1–21.
From:蔡溯林