文章题目:Chitinolytic Streptomyces griseorubens E44G enhances the biocontrol efficacy against Fusarium wilt disease of tomato
文章链接:http://apps.webofknowledge.com/full_record.do?product=UA&search_mode=GeneralSearch&qid=1&SID=5CjEPGpitbbFqFn8oCp&page=1&doc=1
具有几丁质分解能力的灰略红链霉菌 E44G增强番茄枯萎病的生防效果
Phytoparasitica (2017) IF 1.2
E44G是一种从沙特阿拉伯(炎热,干旱的气候区域)的栽培土壤中分离的细菌。在体外,针对植物病原真菌Fusarium oxysporum f.SP.lycopersici(番茄枯萎病的病原体)评估了S. griseorubens E44G的抗真菌潜力。当使用胶体几丁质琼脂平板法时,证明了S. griseorubensE44G的几丁质分解活性。使用凝胶过滤色谱法纯化分子量为45kDa的热稳定性几丁质酶。在60℃和pH 5.5下获得最佳活性。纯化的酶对植物病原真菌F.oxysporum显示出非常显着的活性。在田间评估中,与未感染的未经处理的番茄植物相比,应用两种S. griseorubensE44G处理显着增加了所有测试的生长和产量参数并降低了疾病严重性,表明其可能作为生物防治剂。
枯萎病,由尖孢镰刀菌(Fusarium oxysporum f.SP.lycopersici)引起。是世界上最严重的番茄病之一。它会对植物造成严重损害,导致番茄产量的高经济损失。
几丁质酶是糖基水解酶,其催化β-1,4-键的水解切割。几丁质酶具有重要意义,因为它们具有广泛的生物加工和生物技术方面。在农业中,它们在植物致病真菌的生物控制中具有潜在的应用。它们的抗真菌活性归因于对几丁质的水解作用,几丁质构成真菌的主要结构组分。已知几丁质酶由几种生物产生,包括细菌和放线菌,真菌,植物,无脊椎动物和脊椎动物。链霉菌属菌株被认为是土壤中几丁质酶的主要生产者。由于它们的抗真菌活性,链霉菌属菌株已被广泛研究作为针对不同植物病原真菌的生物控制剂。在我们以前的研究中,筛选了几种从沙特阿拉伯土壤中分离出的放线菌菌株的抗真菌潜力。产生几丁质酶的有希望的分离物之一被鉴定为S. griseorubens E44G。
此次研究涉及
1)体外筛选S. griseorubensE44G对抗植物病原真菌F.oxysporum的抗真菌潜力,
2)来自S. griseorubensE44G的几丁质酶的纯化和表征
3)评估其应用的可能性在田间条件下用作番茄枯萎病的生防剂。
材料和方法
从番茄植物中分离出真菌病原体。将病原真菌在含有马铃薯葡萄糖琼脂培养基(PDA)的平板上培养,并在25±2℃温育72小时,然后使用单孢子技术纯化。
在先前的研究中,从从沙特阿拉伯收集的栽培土壤样品中分离使用的细菌菌株。根据形态学,生物化学和生理学特征鉴定分离的细菌(S.griseorubens E44G),并使用16S rRNA基因进行分子确认。在4℃下将该放线菌的原种培养物保持在淀粉硝酸盐/ NaCl斜面培养基上。
方法:
筛选抗真菌潜力
制备淀粉硝酸盐琼脂(高氏一号)平板(3个重复),其接种了作为孢子悬浮液的F.oxysporum,然后固化。在平板的中心,放置来自新生长的S.griseorubens E44G培养物的圆盘(直径6mm)并在31℃温育。在孵育3天后,以透明区(如果有的话)的形式记录抗真菌潜力。
几丁质分解潜力的测定:
使用含有0.1%胶体几丁质和1.5%琼脂的胶体几丁质琼脂平板评估壳多糖溶解潜力,并在31℃温育。 72小时后,观察到在接种点周围形成清晰区域。
胶体几丁质的制备根据Khan等人的方法制备胶体几丁质。在250毫升浓盐酸中加入20g几丁质粉末,在5℃下连续搅拌溶解1小时。为了除去过量的酸,将甲壳质用蒸馏水洗涤5次,然后用滤纸过滤。将收集的胶体几丁质在5℃下保持糊状。
几丁质酶活性测定
如Khan等人所述,估计培养物上清液和所有纯化步骤中的几丁质酶活性。几丁质酶活性的反应溶液含有50mM乙酸盐缓冲液; 1 mL,5%胶体几丁质;在pH 5和粗酶下1 mL; 1mL,并在50℃下孵育1小时。为了终止反应,煮沸15分钟,然后将228 Phytoparasitica(2017)45:227-237溶液以5000rpm离心15分钟。基于释放的GlcNAc(Monreal和Reese 1969)的浓度,使用分光光度计(UV-260,Shimadzu,Japan)在540nm处通过分光光度法测量几丁质酶活性。在所述测定条件下产生1mM.min -1的GlcNAc的酶量被认为是几丁质酶活性的单位
蛋白质含量的测定:
蛋白质含量的测定如Bradford(1976)所述进行。反应混合物由酶样品组成; 10μmL-1,Bradford溶液; 990μ.mL-1(考马斯亮蓝G-250,Serva,海德堡,德国,目录号17524.01)并用蒸馏水调节至200mL的体积。 3分钟后,使用分光光度计在595nm记录吸光度。对于空白样品,使用0.9%NaCl(10μmL-1)。为了制备标准曲线,使用牛血清白蛋白(Sigma,USA,目录号A9056)。
几丁质酶的纯化:
1.酶沉淀
使用硫酸铵(70%),在上清液中沉淀蛋白质含量。使用磁力搅拌器连续搅拌粗酶,并在5℃下保持12小时,然后以8000rpm离心20分钟,然后收集沉淀并重悬于乙酸钠缓冲液(pH5)中并透析过夜。
2.凝胶过滤色谱
通过过滤器(0.45μm孔径)过滤重悬的蛋白质沉淀,然后使用pH5(蛋白质带负电荷)的50mM乙酸钠缓冲液加载到离子交换层析Mono Q柱(带正电)(GE Healthcare Life science,Piscataway,NJ,USA)中。级分(收集3mL),流速为1mL.min-1。
在评估所收集的几丁质酶的级分后,合并阳性级分并装入Sephadex G-100柱(Sigma,USA,目录号G100120)中,使用pH5的50mM乙酸钠缓冲液和NaCl(100) mM)在0.5mL.min-1流速下。
几丁质酶分子量测定:
十二烷基硫酸钠聚丙烯酰胺凝胶电泳(SDS-PAGE)
如Laemmli(1970)所述,将纯化的几丁质酶进行聚丙烯酰胺凝胶电泳。然后将凝胶染色过夜(考马斯亮蓝G-250; 0.25g.mL -1,甲醇; 25%和乙酸; 10%),然后在25%甲醇和乙酸中脱色12小时。标准蛋白质用作标记物。
几丁质酶的稳定性:
pH对几丁质酶活性和稳定性的影响
通过在0.5个pH单位间隔下改变反应混合物的pH值,在3和9.5之间来估计几丁质酶的最佳pH。为了调节pH值,使用以下缓冲液(pH8-9的Tris-HCl缓冲液(50mM),pH7的磷酸盐缓冲液(50mM)和pH3-6的乙酸盐缓冲液(50mM)。为了估计酶活性,将2mL缓冲液加入酶溶液(100mL)中并在36℃下孵育31分钟。通过在相同温度下在不同pH下孵育酶溶液而没有底物1小时来估计几丁质酶的pH稳定性。
温度对几丁质酶活性和稳定性的影响
为了确定几丁质酶活性的最佳温度,将反应混合物在最佳pH(5.5)下在不同温度(20-80℃)下温育35分钟。纯化酶的热稳定性通过估计在先前温度pH5.5下孵育1小时后没有底物的残留酶活性来确定。
纯化酶的抗真菌潜力
使用琼脂孔扩散法估计纯化酶的抗真菌潜力(Taechowisan等人,2003)。制备测试真菌的孢子悬浮液,并调节浓度为10^5孢子.mL-1,并在凝固前接种到PDA平板上。使用无菌软木钻孔器将孔(直径7mm)制成琼脂板。将酶溶液(50μL)加入孔中,并将热煮的酶溶液用作对照,一式三份重复。在25±2℃温育3天后估计酶活性作为抑制区的直径。
田间评估该实验
在具有已知枯萎病历史的田地中在自然感染的土壤条件下进行的。实验单元(plot)是3m×2m的区域,包含6行。在同一排(小巷)的地块之间跳过一米。在街区之间跳过过道(50厘米宽的拖拉机道路)。将地块犁好,除去所有杂草,并将土壤平整。按建议施用氮肥和磷肥。移植30天大的幼苗。该实验以随机完整区组设计进行。处理可归纳如下:仅病原体(P),病原体+化学杀真菌剂(PF),病原体+ S. griseorubensE44G(PS)的孢子悬浮液,以及病原体+S. griseorubensE44G无细胞培养滤液(PA))。在移植前,将番茄幼苗的根浸泡在浓度为10^6孢子的E44G的孢子悬浮液中,无细胞培养滤液和化学杀真菌剂(Rhizolex-T)浸泡1小时。对于阴性感染对照处理,在移栽前将番茄幼苗的根浸入淡水中。每种处理使用四个plot。在移植后10,25和40天重复应用生物防治处理作为土壤浇灌。
根据Epp(1987)制定的标准评估疾病严重程度。移植后6周,用整个根系小心地取出每种处理的15株植物,洗涤并评估生长参数(植物高度,植物新鲜和干重以及叶子数)。在生理成熟时,收获来自每个小区的番茄果实并分别称重以确定果实产量。
统计分析
分析软件BCoStat 6.311用于所有统计分析。使用Duncan的New Multiple Range Test测定治疗方法之间的显着差异。
结果和讨论
S.griseorubens E44G的抗真菌活性在体外,针对植物病原真菌F.oxysporum评估了S.griseorubens E44G的抗真菌潜力。孵育72小时后,以24mm的抑制区形式抑制F.oxysporum的生长(图1)。该结果与Alekhya和Gopalakrishnan(2014)的结果一致,筛选了27种放线菌分离株对抗真菌病原体的拮抗作用,并记录了8种分离物在双重菌株培养试验中表现出广谱拮抗活性。这种拮抗性质可归因于过度寄生,抗菌物质或水解酶的产生。
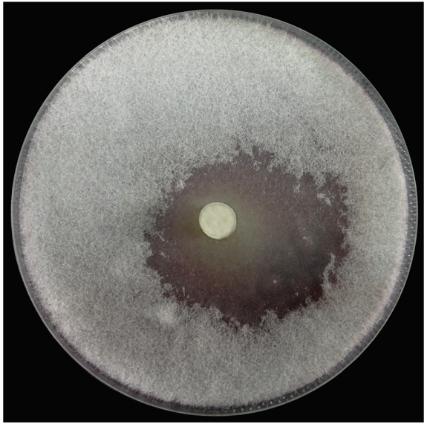 (图1)
(图1)
筛选几丁质酶活性
在测试的分离物的圆盘周围记录抑制区,表明几丁质水解。该结果证明了壳聚糖活性有助于S. griseorubensE44G的拮抗性质。
链霉菌属菌株被认为是土壤中主要的几丁质酶生产者。已经报道了来自不同链霉菌属菌株的不同程度的几种几丁质酶,包括S. rimosus,S.griseus,S.plicatus,S.lividans和S. halstedii。
几丁质酶的纯化通过蛋白质沉淀和凝胶过滤色谱法进行
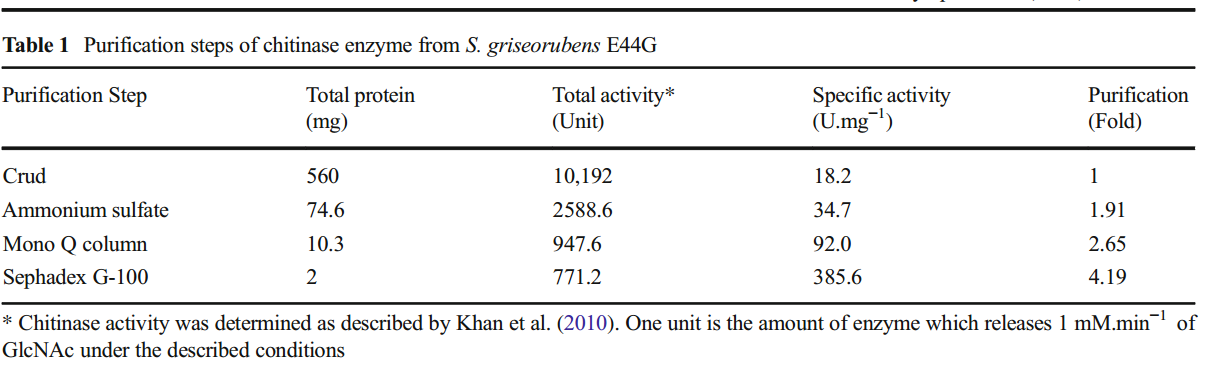
S.griseorubens E44G的细胞外几丁质酶的纯化。在每个阶段,评估蛋白质含量和酶活性(表1)。粗酶显示出18.2U.mg-1的比活性,而硫酸铵沉淀的酶显示出34.7U.mg-1的比活性,纯化倍数为1.91。在70%的硫酸铵下获得几丁质酶的最大沉淀,与粗酶和其他浓度相比显示出最高的几丁质酶活性(数据未显示)。
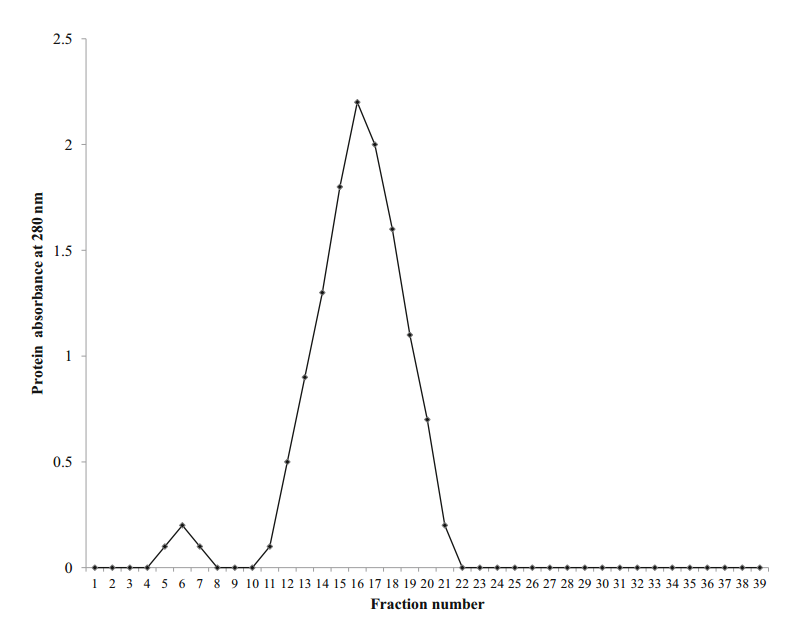
使用Mono Q柱的酶纯化公开了92U.mg-1的比活性,纯化倍数为2.65。在筛选几丁质酶的级分后,将阳性管的内容物加载到sephadex G-100柱中。几丁质酶溶液的洗脱曲线如图2所示。
在收集的馏分中,活性部分(NO.16)显示最高比活性为385.6U.mg-1,纯化倍数为4.19。 Kavitha和Vijaylakshmi(2011)在sephadex G-100纯化后记录了来自S. tendae TK-VL 333的几丁质酶的纯化倍数为2.96的37.4U.mg-1的比活性。另一方面,根据Rabeeth等人的研究。通过sephadex G-100柱纯化的S.griseus MTCC9723的几丁质酶表达15,500Umg-1的比活性和9.3的纯化倍数。
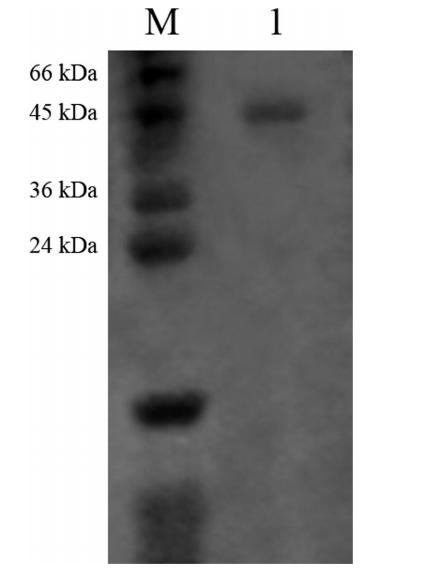
纯化几丁质酶的分子量使用SDS-PAGE分析,证实来自S. griseorubensE44G的纯化几丁质酶的同质性显示~45kDa的单一蛋白质条带(图3)。该发现与先前研究中Narayana和Vijayalakshmi(2009)报道的来自链霉菌属(Streptomyces sp.)链霉菌ANU 627和S. tendae T KVL 333产生的分子量为45kDa的几丁质酶的结果一致。先前报道过的几丁质酶分子量范围在20和90kDa之间。
与植物几丁质酶类似,细菌几丁质酶的分子量范围为20-65kDa,但小于昆虫几丁质酶(40-85kDa)。据报道,从不同链霉菌获得的几丁质酶具有不同的分子量,如来自链霉菌属(Streptomyces sp.TH-11)的29kDa,来自S.griseus MTCC 9723的32kDa,34kDa,来自链霉菌属ANU 6277的45kDa,来自S. violascens NRRL B2700的65 kDa。
几丁质酶的表征
pH对几丁质酶活性和稳定性的影响
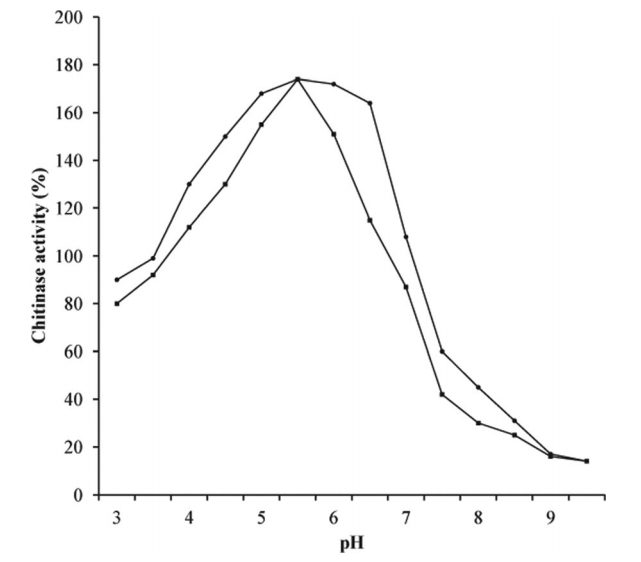
图4中所示的数据表示pH值对纯化的几丁质酶在3-9.5的pH范围内的活性和稳定性的影响。酶活性随着pH值的增加而增加,并且在pH5.5时表现出最大的活性(图4)。 pH值的进一步增加导致几丁质酶活性降低。然而,超过该pH值,几丁质酶活性迅速下降。该酶在pH4-7下稳定,保持至少65%的活性。据报道,许多链霉菌几丁质酶在酸性pH范围内显示出它们的最佳活性。
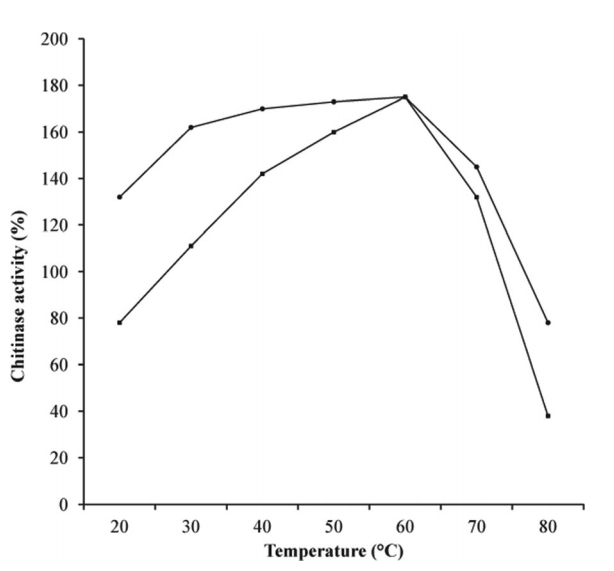
纯化酶的热稳定性图5显示了纯化酶在各种温度下的活性。S. griseorubensE44G的几丁质酶在30-70℃范围内表现出相当大的活性。最佳温度记录为60℃,之后,酶活性随培养温度的进一步升高而降低。几丁质酶在30至60℃的温度范围内保持超过90%的活性,表明该酶在高达60℃的温度下是稳定的。 S. griseorubensE44G的几丁质酶的热稳定性在工业规模上提供了重要的特征。根据我们的结果,Nagpure等人。 (2013)报道了来自S. violaceusniger MTCC 3959的几丁质酶产生的最佳温度为60℃。另一方面,Rabeeth等人记录了40℃作为来自灰色链霉菌MTCC9723的几丁质酶产生的最佳温度,并且在20至50℃的稳定性。
纯化酶的抗真菌活性记录了清晰的抑制区(30mm)(图6),而灭活的酶没有显示出任何抑制作用。产生几丁质酶的生物可以直接使用它们的代谢物或基因用于植物病原体的生物控制。 Mukherjee和Sen(2006)使用来自委内瑞拉P10的纯化几丁质酶记录了对黑曲霉,Helminthosporium sativum和Alternaria alternata的显着生长抑制。 Ghasemi等人获得了相同的结果。
田间评价影响S. griseorubensE44G对番茄生长参数和疾病严重程度的影响
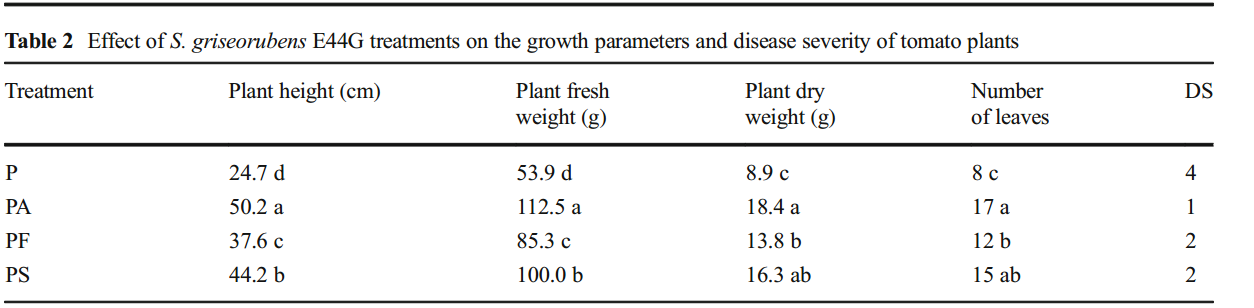
表2列出了生物防治处理对感染枯萎病的番茄植株生长参数和疾病严重程度的影响。
与感染的未处理的番茄植物相比,用E44G的无细胞培养滤液处理的感染植物显示出所有测试的生长参数的显着增加。感染植物的疾病严重程度显着。链霉菌菌株与未处理的感染植物相比,用S. griseorubensE44G的无细胞培养滤液处理时降低。用S. griseorubensE44G的孢子悬浮液处理的感染植物排名第二最佳处理,然后用杀真菌剂处理。
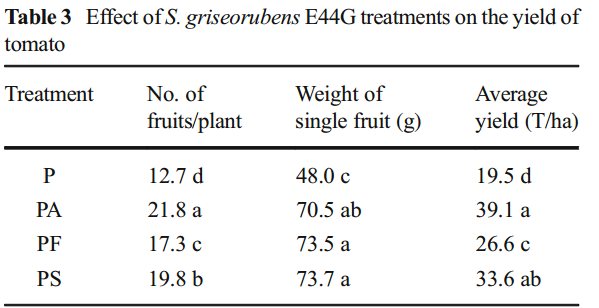
S. griseorubensE44G处理对番茄产量的影响表3中显示的数据显示了S. griseorubensE44G处理对番茄产量的影响。
枯萎病的感染导致感染的番茄植物的产量参数显着降低。然而,与未处理的感染植物和用化学杀真菌剂处理的植物相比,用滤液处理处理的感染植物的产量显着增加。用S. griseorubensE44G的孢子悬浮液处理感染的植物是第二好的处理。
用滤液处理处理的感染植物的产量增加可归因于滤液组分的生长调节作用。
这些结果与Yandigeri(2015)等人获得的结果一致,在番茄植物上使用壳多糖溶解放线菌Streptomyces vinaceusdrappus S5 MW2。已经报道了壳多糖溶解放线菌在植物病害的生物控制中的潜力。除了抗生素和竞争之外,水解酶作用似乎也有助于它们的拮抗行为。
单独或与生物控制剂组合的几丁质的抗性诱导效应已被认识到。这些重要特征除了高温生境下S. griseorubensE44G的存活外,使该分离物成为一种良好的生物防治剂。
结论
在本研究中,观察了S. griseorubensE44G对F.oxysporum的体外抗真菌潜力。显示壳聚糖活性有助于S. griseorubensE44G的拮抗行为。使用凝胶过滤色谱法纯化分子量为45kDa的热稳定性几丁质酶。在60℃和pH 5.5下获得最佳活性。纯化的酶对植物病原真菌F.oxysporum显示出非常显着的活性。在田间评价中,与未经处理的感染番茄植物相比,应用S. griseorubensE44G处理显着增加了所有测试的生长和产量参数并降低了疾病严重性,表明可能作为生物防治剂。另一方面,S.griseorubens E44G的几丁质酶的热稳定性可能在工业规模上提供重要的特征。
from方悠