Highlights
Soil fauna generally increased extracellular enzyme production by basidiomycetes.
Grazing effects were species-specific; macrofauna induced the greatest responses.
Two fungal functional groups showed opposite responses to grazing.
Enzymatic responses occurred in grazed and un-grazed regions of mycelium.
Enzymes involved in carbon and phosphorous cycling were most frequently affected.
Introduction
Extracellular enzymes produced by heterotrophic microorganisms in the soil are responsible for the decomposition of organic compounds.
Basidiomycete fungi are the primary decomposer agents in temperate wooded ecosystems and contribute extensively to extracellular enzyme activity and nutrient mineralisation within soils.
Growth and development of basidiomycete mycelia is influenced by soildwelling invertebrate grazers with potential implications for fungal activity and ecosystem functioning.
Purpose
To date, no study has attempted to quantify or compare the impacts of individual invertebrate species on enzyme production by basidiomycetes.Attempted to quantify or compare the impacts of individual invertebrate species on enzyme production by basidiomycetes.
Materials and methods
Invertebrate : Myriapoda(Blaniulus guttulatus) Isopoda(Oniscus asellus) Collembola(Folsomia candida) Nematoda(Panagrellus redivivus)
Basidiomycete : Phanerochaete velutina, Resinicium bicolor and Hypholoma fasciculare DD3 Hypholoma fasciculare JH
Fungal culturing and inoculation
2%malt extract agar
Freshly-felled beech wood (Fagus sylvatica) was cut into blocks (2*2*1 cm)(sterilised)
Petri dishes were sealed with Nescofilm and incubated in the dark for three months at
21℃ to allow fungal colonization .
Soil, collected to a depth of 20 cm from deciduous woodland was sieved on site through a 10mmmesh and air-dried in plastic trays. Dried soil was sieved through 2 mm mesh and frozen for 24 h at 18℃to kill any remaining soil fauna. Prior to use, soil was re-wetted with 340 ml DH2O kg soil-1.200 g wet soil was then smoothed and compacted to a depth of 5 mm within 24*24 cm bio-assay dishes.
Preparation of 125 trays allowed for:
(i) five replicates of each interaction (interaction trays);
(ii) five control trays of each fungus growing without grazers (fungus controls);
(iii) five controls of each invertebrate species with no fungus (invertebrate controls);
(iv) five control trays with no organisms added (soil controls).
Fungus-colonised wood blocks were cleaned of surface mycelia and placed into the centre of soil trays. Oncemycelia in the trays for each fungal strain had reached approximately 8 cm diameter any visible fungal contamination was removed, and invertebrates were introduced around the basidiomycete mycelium onto un-colonised soil.
After invertebrate addition, trays were sealed in polythene bags to reduce water loss and microbial contamination. Bags were incubated at 21℃and 70% relative humidity.
Soil samples for enzyme analysis
Soil samples were extracted 10 d after invertebrate addition (when substantial grazing had been observed in all trays). Trays were cut into a grid of squares (1 cm3) with each square representing one sample. 30 samples were taken from each interaction and fungus control tray: of the samples, 10 contained soil colonized by old mycelium (2 cm from the wood block) (OM), 10 had soil colonised by young mycelium (1 cm from hyphal tips) (YM) and 10 were from un-colonised soil (S). Ten samples each of un-colonised soil were also extracted randomly from invertebrate and soil control trays. Samples from the same regions of soil from each tray were mixed and stored in a freezer (-18℃).
Enzyme assays
Extracellular enzyme activity was determined in multiwell plates using a modified method of Vepsäläinen et al. (2001).
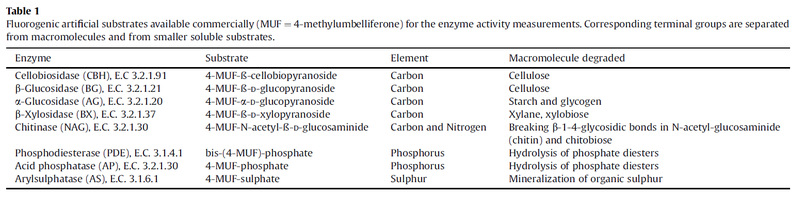
Image capture and analysis
Soil trays were photographed using a Nikon Coolpix 57000 camera, mounted on a stand at a height of 40.5 cm. Images of each tray were captured on the first and final day of each interaction. Images were analysed using IMAGEJ (National Institutes of Health, USA).
Statistical analysis
R version 2.10.1
A Gaussian General Linear Model (GLM) was also performed (with coverage, fungus and invertebrate treatment as factors) to test for associations between enzyme production and hyphal coverage in different grazing treatments.
Results
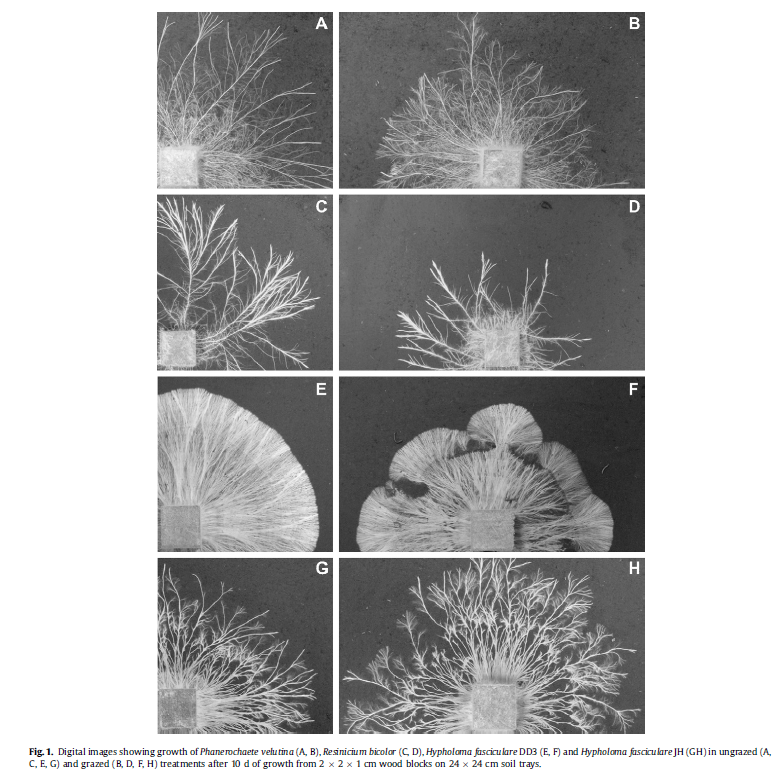
Mycelial distribution varied between fungi. Both H. fasciculare DD3 and Phanerochaete velutina produced densely aggregated mycelia, covering the majority of soil within the diameter of their respective networks. R. bicolor produced thick, loosely aggregated mycelia, displaying a more explorative foraging strategy. H. fasciculare JH morphology was an intermediate between these two foraging strategies, producing thick mycelia with branched tips.
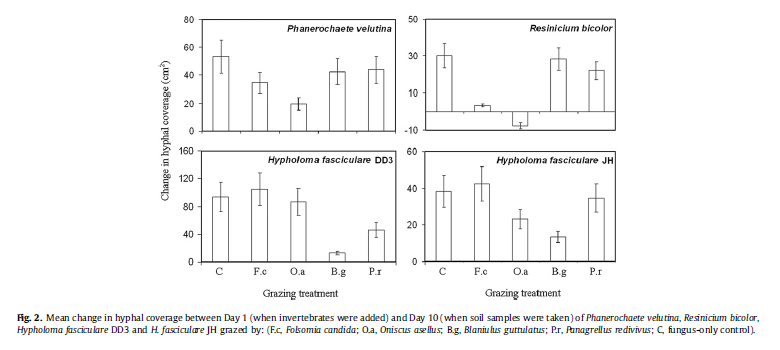
Fungi also varied in their susceptibility to grazers. Both R. bicolor and Phanerochaete velutina were grazed heavily by Oniscus asellus and F. candida. Oniscus asellus grazing significantly reduced hyphal coverage of Phanerochaete velutina and R. bicolor. F. candida also reduced Phanerochaete velutina and R. bicolor but to a lesser extent than Oniscus asellus (Fig. 2). The fungi, however, responded very differently to grazing.
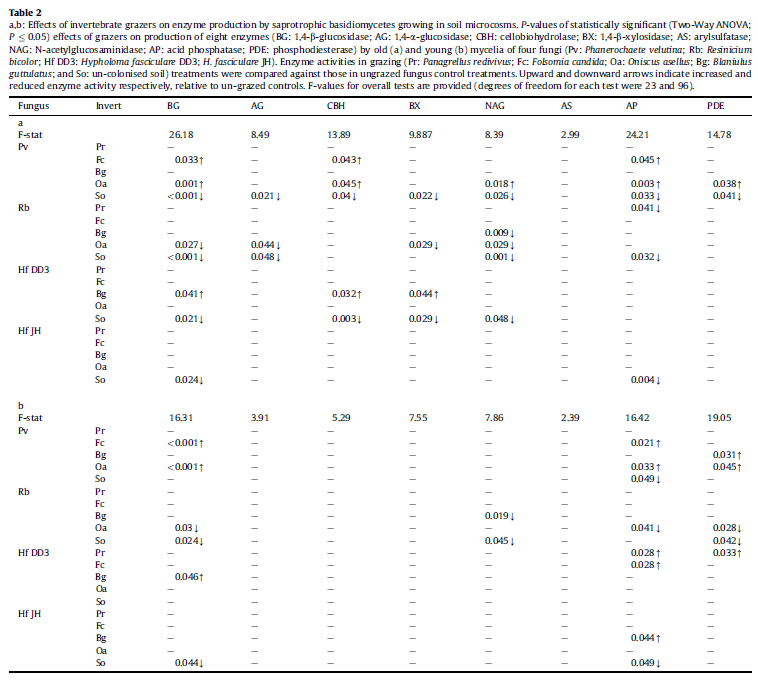
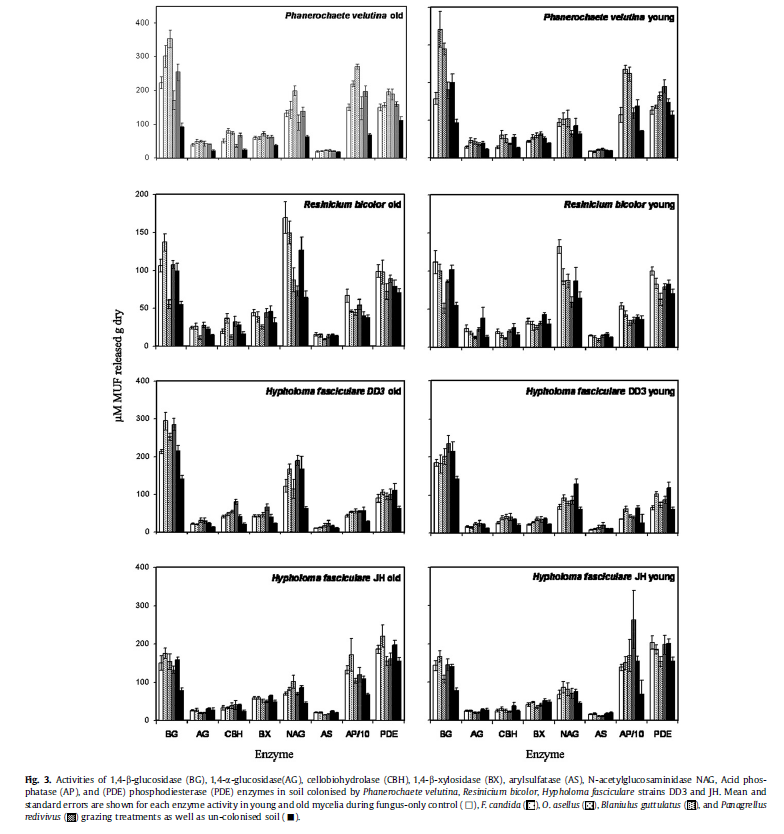
For each enzyme, activity was not significantly (GLM; P > 0.05) affected by hyphal coverage in any treatment. Recorded changes in enzyme production by fungduring grazing were, therefore, not due to changes in fungal biomas All recorded changes in soil colonised by Phanerochaete velutina and H. fasciculare DD3 represented increases in enzyme activity. Effects ranged from being insignificant (AS) to being almost twofold (BG, AP) greater than in ungrazed fungus controls (Fig. 3). In contrast, enzyme activity under R. bicolor cords was always reduced by invertebrate grazing. Enzyme production by H. fasciculare JH was generally unaffected by grazing; only AP activity increased during millipede grazing (Table 2).
For each fungal strain, the invertebrate species which were responsible for the greatest reduction in hyphal coverage also exerted the greatest influences on enzyme activity. Oniscus asellus caused the greatest number of significant effects to Phanerochaete velutina and R. bicolor, significantly affecting the activities of five and six enzymes, respectively (Fig. 3). Folsomia candida significantly increased BG, CBH and AP activities in soil colonised by Phanerochaete velutina and AP activity under H. fasciculare DD3 mycelia (Table 2). B. guttulatus exerted the greatest number of changes to enzyme activity under both H. fasciculare strains. Millipedes stimulated the production of BG, BX and CBH by H. fasciculare DD3mycelia and were the only species to influence the activity of any enzyme in H. fasciculare JH trays (Fig. 3).
Most enzymatic responses to grazing were similar in young and old regions of mycelia (Fig. 3). Activities of all enzymes except AS were significantly (P 0.05) affected by some grazing treatments (Fig. 3). Certain enzymes were, however, more frequently affected.
Although all grazers influenced mycelial enzyme production, at least to some extent,impacts varied dramatically between invertebrate species. This may be related to grazing intensity. For example, the greatest number of changes to Phanerochaete velutina and H. fasciculare DD3 enzyme production were induced by woodlice (Oniscus asellus) and millipedes (B. guttulatus), respectively-the same invertebrate species that reduced hyphal coverage of these fungi most markedly in this and previous studies. Invertebrates, therefore, have species-specific impacts on fungal enzyme production; spatial organisation and community compositions of soil fauna will have direct consequences for nutrient mineralisation within forest soil.
From a fungal perspective, impacts of grazing were less variable; enzyme activities under H. fasciculare DD3 and Phanerochaete velutina mycelia always increased during grazing while activities under R. bicolor were always reduced. The striking contrast between changes in enzyme activity under different fungi, however, suggest that enzymatic changes were the result of specific fungal responses to grazing rather than mechanistic damage. Therefore effects of mycophagous soil fauna on fungal-mediated nutrient mineralisation will vary between habitats, depending on the dominant fungal species present.
The enzymatic response of a fungus may relate to its growth and activity during invertebrate invasion.
The consistent changes to enzymes involved in cellulose decomposition and phosphate acquisition during grazing suggest that mycophagous fauna may be particularly influential in the cycling of carbon and phosphorus in woodland ecosystems.
Conclusion
Grazing was associated with an increase in enzyme production but individual effects varied between invertebrate species. Opposing enzymatic responses of different fungal functional groups mean that the impacts of grazing soil fauna vary between fungal communities.
From chenyunru