Title:Lignin lags, leads, or limits the decomposition of litter and soil organic carbon
Doi: 10.1002/ECY.3113
Abstract:Lignin’s role in litter and soil organic carbon (SOC) decomposition remains contentious. Lignin decomposition was traditionally thought to increase during midstage litter decomposition, when cellulose occlusion by lignin began to limit mass loss. Alternatively, lignin decomposition could be greatest in fresh litter as a consequence of co‐metabolism, and lignin might decompose faster than bulk SOC. To test these competing hypotheses, we incubated 10 forest soils with C4 grass litter (amended with 13C‐labeled or unlabeled lignin) over 2 yr and measured soil respiration and its isotope composition. Early lignin decomposition was greatest in 5 of 10 soils, consistent with the co‐metabolism hypothesis. However, lignin decomposition peaked 6–24 months later in the other five soils, consistent with the substrate‐limitation hypothesis; these soils were highly acidic. Rates of lignin, litter, and SOC decomposition tended to converge over time. Cumulative lignin decomposition was never greater than SOC decomposition; lignin decomposition was significantly lower than SOC decomposition in six soils. Net nitrogen mineralization predicted lignin decomposition ratios relative to litter and SOC. Although the onset of lignin decomposition can indeed be rapid, lignin still presents a likely bottleneck in litter and SOC decomposition, meriting a reconsideration of lignin’s role in modern decomposition paradigms.
Materials and Methods
Collected surface mineral soils (0–10 cm, surface litter removed) from 10 North American forests spanning a broad range of edaphic characteristics.
Standing senesced above-ground stem and leaf tissue (litter) from Andropogon gerardii Vitman, a C4 grass.
13C-labeled and unlabeled lignins
Soil incubation experiments
Two treatments were imposed with three replicates for each soil: amendment with 100 mg A. gerardii litter + natural abundance lignin, or with 100 mg A. gerardii litter + 13Cβ-labeled lignin.
Samples were incubated in 50 ml centrifuge tubes placed inside 946-ml glass jars.
Sampling intervals increased from weekly to biweekly to monthly to bimonthly over the two-year incubation to ensure sufficient CO2 accumulation for precise analysis.
Data analysis
Results
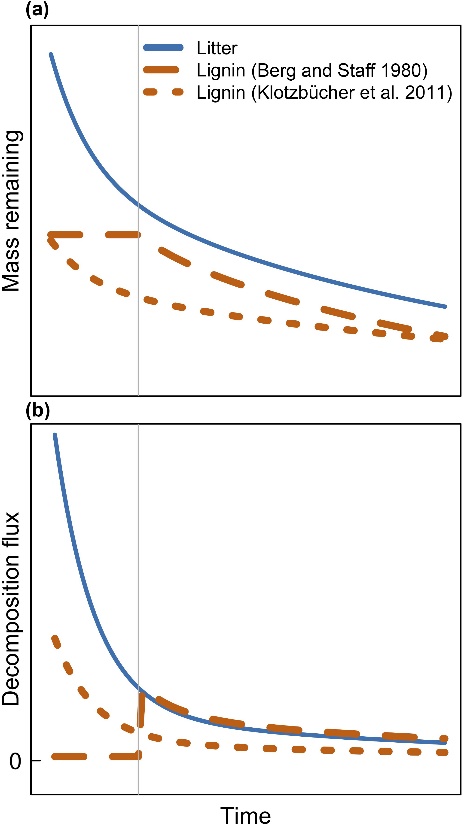
Fig. 1 Conceptual models comparing bulk litter and lignin decomposition over time as (a) mass remaining or (b) instantaneous fluxes. The vertical gray line indicates the transition from early‐ to late‐stage decomposition. In the Berg and Staaf (1980) model, lignin decomposition is low until unprotected litter C has been depleted but controls mass loss thereafter. In the Klotzbücher et al. (2011) model, lignin decomposition is greatest during early litter decomposition and subsequently declines relative to bulk litter decomposition.
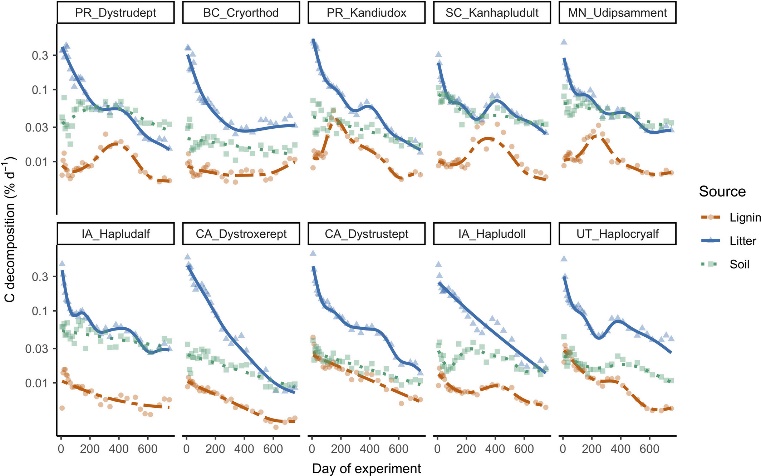
Fig.2 Decomposition rates of lignin Cβ (orange circles), litter C (blue triangles), and soil organic carbon (green squares) for each soil over time, expressed relative to initial C mass in each pool (note the logarithmic y‐axis scale). Lines represent generalized additive mixed model fits. The first two capital letters of each panel denote the state/province of soil origin, followed by the great group in the U.S. Department of Agriculture soil taxonomy.
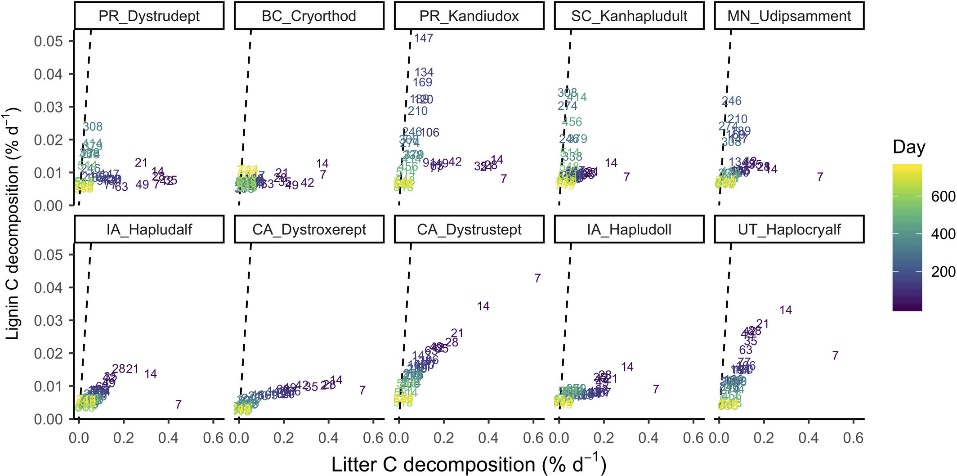
Fig.3 Pairwise relationships between instantaneous decomposition rates of litter C and lignin Cβ for each soil. Numbers and shading denote the date of measurement. The dashed line indicates a 1:1 relationship.
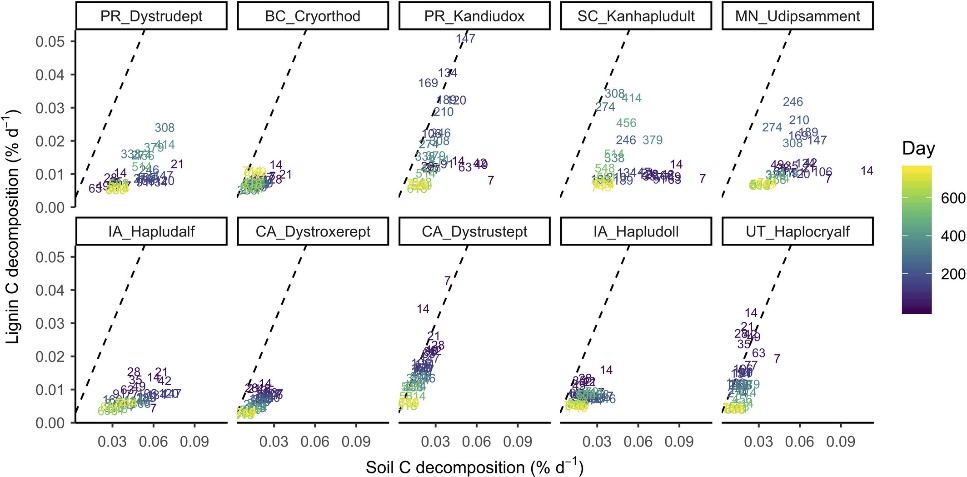
Fig.4 Pairwise relationships between instantaneous decomposition rates of SOC and lignin Cβ for each soil. Numbers and shading denote the date of measurement. The dashed line indicates a 1:1 relationship.
Discussion
Our findings reconcile aspects of competing conceptual models that have been used to predict relationships among the decomposition of lignin, litter, and SOC. We found partial support for both alternatives of our first hypothesis: in half of the soils examined, lignin decomposition was greatest early in the experiment and generally decreased thereafter, consistent with the hypothesis that increased labile C supply drives co‐metabolic decomposition of lignin during early decomposition (Klotzbücher et al. 2011). However, in the other five soils, lignin decomposition subsequently increased and exceeded initial values after many months. This is consistent with the substrate limitation hypothesis, whereby lignin decomposition increases as carbohydrate availability decreases to enable access to protected substrates (Berg and Staaf 1980). Even across these contrasting groups of soils, however, we found common threads of evidence supporting both co‐metabolism and substrate limitation.
What ecological factors, then, might predict differences in the temporal patterns of lignin decomposition rates and cumulative lignin losses among soils? Availability of N might play an important role, as N enrichment has been linked to suppression of lignin decomposition and accumulation of SOC at local and global scales .We also found circumstantial evidence for the role of soil pH as a control on lignin‐degrading organisms.
from Heweihua