Title: Environmental stress response limits microbial necromass contributions to soil organic carbon
http://dx.doi.org/10.1016/j.soilbio.2015.03.002
Abstract:
The majority of dead organic material enters the soil carbon pool following initial incorporation into microbial biomass. The decomposition of microbial necromass carbon (C) is, therefore, an important process governing the balance between terrestrial and atmospheric C pools. We tested how abiotic stress (drought), biotic interactions (invertebrate grazing) and physical disturbance influence the biochemistry (C:N ratio and calcium oxalate production) of living fungal cells, and the subsequent stabilization of fungal-derived C after senescence. We traced the fate of13C-labeled necromass from ‘stressed’ and‘unstressed’ fungi into living soil microbes, dissolved organic carbon (DOC), total soil carbon and respired CO2. All stressors stimulated the production of calcium oxalate crystals and enhanced the C:N ratios of living fungal mycelia, leading to the formation of ‘recalcitrant’ necromass. Although we were unable to detect consistent effects of stress on the mineralization rates of fungal necromass, a greater proportion of the non-stressed (labile) fungal necromass C was stabilised in soil. Our finding is consistent with the emerging understanding that recalcitrant material is entirely decomposed within soil, but incorporated less efficiently into living microbial biomass and, ultimately, into stable SOC.
Results:
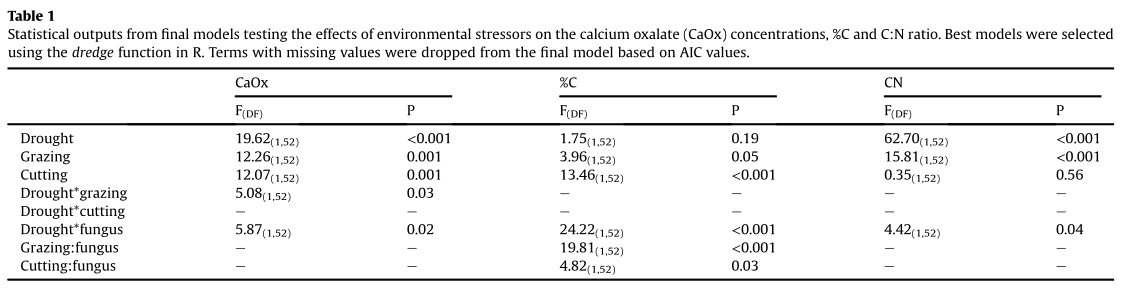

Fig. 1. Electron scanning microscope images of mycelial cords of calcium oxalate crystal production on the cords of Resinicium bicolor (a, c, e) and Phanerochaete velutina (b, d, f)
following growth under optimal conditions (a, b), isopod grazing (c, d) and drought (e, f) Images show that the density and size of crystals accumulated increase during stress.

Fig. 2. Effects of abiotic, biotic and mechanical stress on the calcium oxalate production (a, b) and C:N ratio (c, d) by Resinicium bicolor (a, c) and Phanerochaete velutina (b, d)
growing in compacted soil microcosms. White and gray bars represent fungi grown under optimal (-0.006 MPa) and drought (-0.06 MPa) conditions, respectively. Different letters
above bars refer to significantly (2 way ANOVA: P > 0.05) differences between treatments.
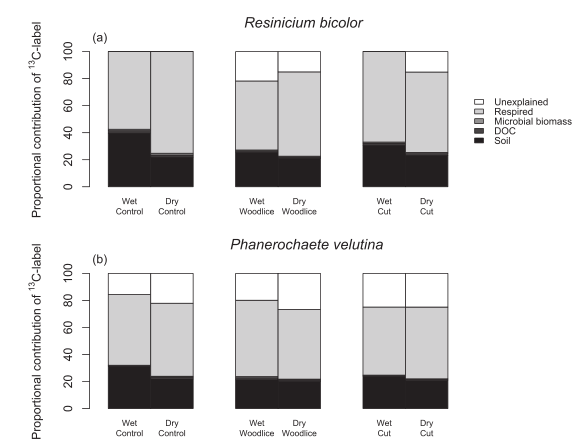
Fig. 3. Proportional contribution of13C-labeled Resinicium bicolor (a) and Phanerochaete velutina (b) to stabilized soil C, dissolved organic C, microbial biomass C and mineralized
(respired) C within decomposition assays. White sections represent un-detected C. It is likely that this excess13C was mineralized between day 2 and 14, and not detected with our
sampling regime.
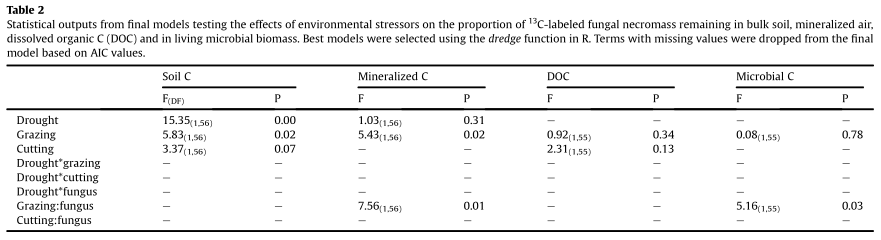
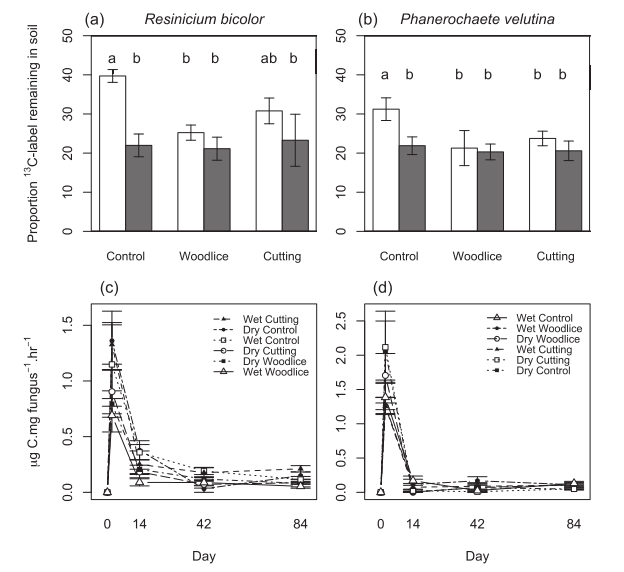
Fig. 4. Effects of abiotic, biotic and mechanical stress on the percentage of initial Resinicium bicolor (a) and Phanerochaete velutina (b) necromass stabilized within soil. White and
gray bars represent the contributions of fungi grown under optimal (-0.006 MPa) and drought (-0.06 MPa) conditions, respectively. Also shows the mineralization rates of13C-
labeled Resinicium bicolor (c) and Phanerochaete velutina (d) over 84 days following addition of necromass to soil microcosms on Day 1. Although initial mineralization rates of the
most labile material tended to be higher than those of stressed necromass, more13C remained in the soil after 84 days.
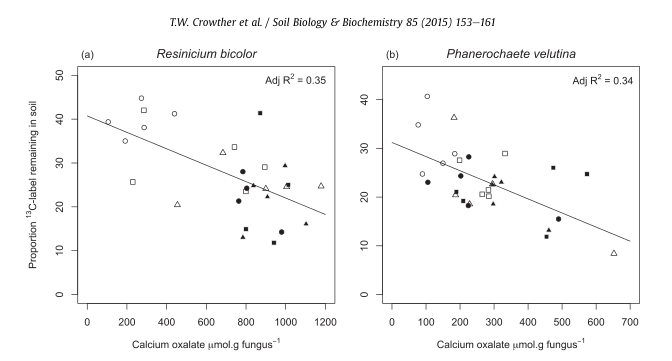
Fig. 5. Linear relationships between fungal calcium oxalate production and proportional necromass contributions to stabilized soil C. The negative effect of calcium oxalate
production on initial SOC formation was equivalent between Resinicium bicolor (a) and Phanerochaete velutina (b). White and black symbols represent the contributions of fungi
grown under optimal (-0.006 MPa) and drought (-0.06 MPa) conditions, respectively. Symbol shapes indicate different treatments (control: circles, grazing: triangles, and cutting:
squares).
Conclusions:
The present study provides a novel mechanism by which environmental stress (biotic interactions, abiotic stress and mechanical disturbance) during the lifetime of an individual microbe, can influence the initial stabilization of its necromass C in soil. Although these stressors have a variety of effects on the partitioning of microbial necromass C between living decomposer biomass, DOC and respired CO2, they consistently reduced fungal contributions to total soil C. It is likely that fungal C contributions to SOC in the present study under-represent those observed under field conditions, as the homogenization of soil might have limited the formation of aggregates that restrict decomposer activity.Furthermore, increases in fungal exudates under stressed conditions might also contribute to changes in soil C stabilization.However, our study excluded other microbial products, focusing on the decomposition of the structural components of microbial cells under controlled laboratory conditions. In doing so, we are able to identify a potential mechanism linking environmental conditions with fungal contributions to soil C. That both fungal species showed similar trends e with reduced stabilization of13C-labeled necromass as stress-induced calcium oxalate production increased-highlights the potential generality in this process, at least across similar taxa. We stress that these results simply highlight the potential for environmental stress responses to influence the stabilization of microbial necromass C in soil. It is now important that we explore the biochemical mechanisms governing this process under field scenarios, due to the potential to explain differences in soil C storage across environments and to improve predictions about the strength of feedbacks between climate change and soil C efflux.
From Chen Yunru