Article 1:
Title: Changes in Volatile Production During the Course of Fungal Mycelial Interactions Between Hypholoma fasciculare and Resinicium bicolor
DOI 10.1007/s10886-006-9209-6
Abstract The mycelia of two wood decay basidiomycete fungi were grown opposing each other across a 1-μm pore membrane supported on the surface of malt broth, contained within a sealable reaction vessel. Production of volatiles during the time course of interaction was followed by collecting head space samples by solid phase microextraction (100 μm polydimethylsiloxane fiber) on five occasions over 25 d following coinoculation of the fungi: 1, 3 (i.e., immediately prior to mycelial contact), 9 (1–2 d after initiation of pigment production by Resinicium bicolor), 17, and 25 d. Ten volatiles were produced during interactions that were not detected in single species controls. In general, most (18) fungal volatiles were sesquiterpenes eluted between 12.5 and 21 min, with a further two eluted at 29.1 and 33.9 min; a benzoic acid methyl ester, a benzyl alcohol, and a quinolinium type compound with a distinctive fragmentation pattern at m/z 203, 204, 206, and 207 were also identified; three volatiles with m/z maxima of 163, 159, and 206–208, respectively, remained unidentified. The results are discussed in relation to possible ecological roles of volatiles.
Hypothesis:
VOCs are produced prior to mycelial contact, initiating an early defense response
VOC production is linked to the production of pigments during interspecific interaction
VOC production increases over the course of the interaction independently of colony size
Interspecific interactions produce additional VOCs, which are otherwise not produced during self interactions
Methods and Materials:
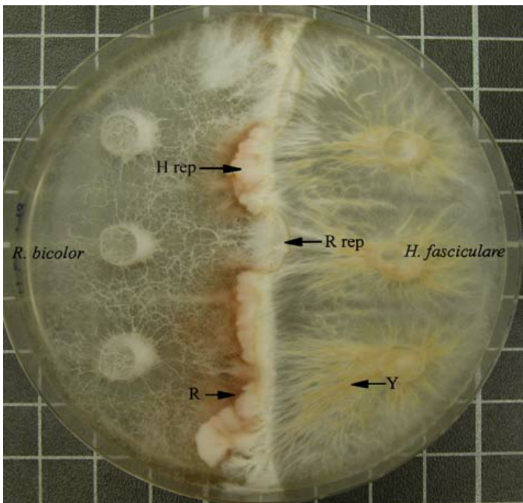
Fig. 1 Resinicium bicolor interacting with Hypholoma fasciculare on 2% malt agar in a 9 cm Petri dish, 18 d after mycelial contact at 20°C. Resinicium bicolor is partially replacing H.fasciculare (R rep) and vice versa(H rep). Note that there is dark red/brown pigment production by R. bicolor (R), and yellow pigment production around cords of H. fasciculare (Y).
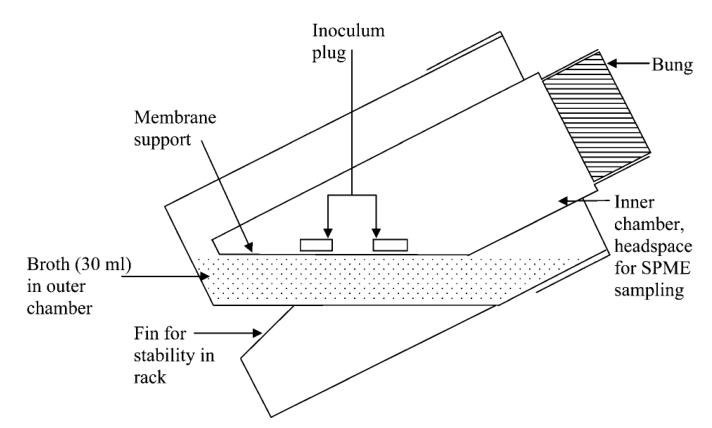
Fig. 2 Diagram of the Reacsyn™ fermentation vessels used to assess production of volatiles and diffusibles when mycelia were interacting and when growing alone
Result:
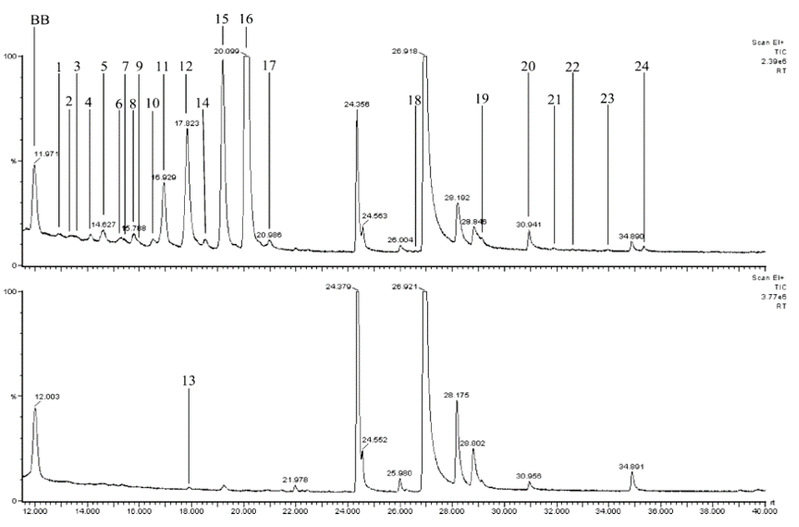
Fig. 3 Representative GC/MS traces indicating fungal metabolites: (a) Hypholoma fasciculare v Resinicium bicolor (HFRB) interaction at 17 d; (b) R. bicolor (RBRB) alone at 17 d indicating position of the peak found in R. bicolor only. Peaks are numbered individually. An additional small peak (25) was found in both treatments at 49.1 min (not shown), BB is dibutylbenzene
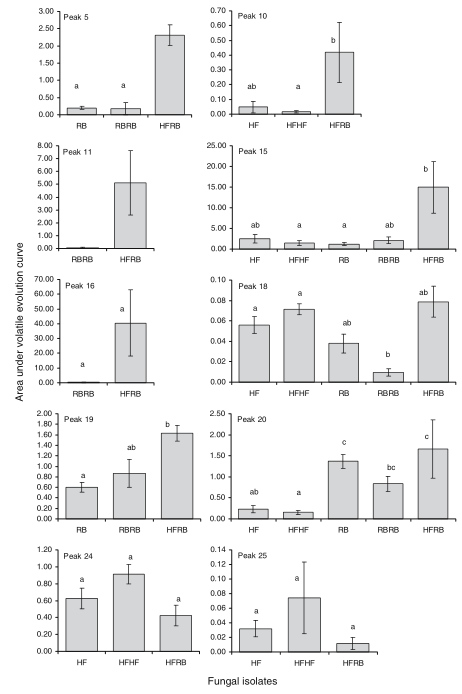
Fig. 4 Evidence of up-regulation of VOCs during interspecific interaction in those volatiles that were also present in single species inoculations: HF, Hypholoma fasciculare inoculated singly; HFHF, H. fasciculare inoculated as two plugs; HFRB, H. fasciculare v Resinicium bicolor; RB, R. bicolor inoculated singly; RBRB, R. bicolor inoculated as two plugs. Values are means (with standard errors) of three replicates of total area under the curve produced over 25 d. For each peak, the same letter indicates no significant difference (P≤ 0.05). Log10 transformation of data were required for peaks 10, 11, 15, 16, and 20. One-way ANOVA with Tukey–Kramer a posteriori test performed on 5, 10, 15, 18–20, and 24. Two-sample t-test was carried out on peaks 11 and 16
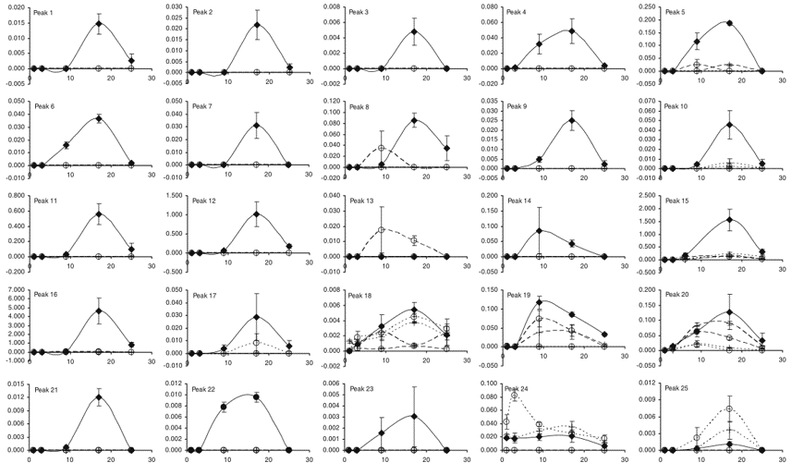
Fig. 5 Production of each volatile (Peaks 1 to 25) in all five treatments over time, values are means (with standard errors) of three replicates. Dotted lines HF, open circles on dotted lines HFHF, dashed lines RB, circles on dashed lines RBRB, solid diamonds HFRB
Discussion:
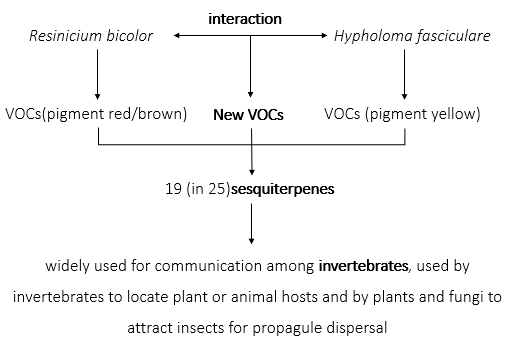
VOC production is linked to the production of pigments during interspecific interaction. VOC production increases over the course of the interaction independently of colony size. Interspecific interactions produce additional VOCs, which are otherwise not produced during self- interactions.
Article 2:
Title: The whiff of decay: Linking volatile production and extracellular enzymes to outcomes of fungal interactions at different temperatures
Download: https://doi.org/10.1016/j.funeco.2019.03.006
Abstract:The terrestrial carbon cycle is largely driven by photosynthetic plants and decomposer organisms that process biomass to CO2. In forest ecosystems, the decomposers are predominantly wood decay fungi, and the response of community structure and activity to increasing global temperatures is likely critical to forest biogeochemical processes. Metabolic products can drive community structure and substrate utilisation, and the role of volatile organic compounds (VOCs), as well as extracellular enzymes, are of particular interest. Pair-wise interactions of a community of basidiomycetes were made under 3 different microclimate conditions that mimic fluctuations in local climate conditions, and the outcome of interactions was assessed in terms of: (1) which fungus won the confrontation or whether it was a draw (deadlock); (2) the production of volatile organic compounds (VOCs) and enzyme activities; and (3) the rate of decomposition. While substrate utilisation and exploitation in terms of decomposition was not affected, community response to changing temperature was underpinned by altered outcomes of interactions and changes to territory occupation, which were reflected by changes in VOC production and extracellular enzyme activity. This study underlines the importance of understanding the impact of community structure on carbon cycling in forest ecosystems under a changing climate.
Hypothesis:
(1) metabolism and interaction outcome are inherently linked, and are affected by environmental conditions;
(2)changes to metabolic strategies for antagonism and resource utilisation reflect alterations to community dynamics.
Materials and methods:
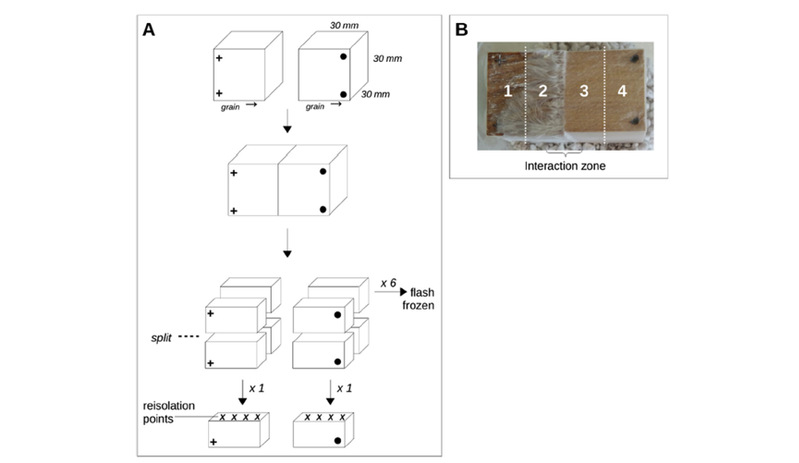
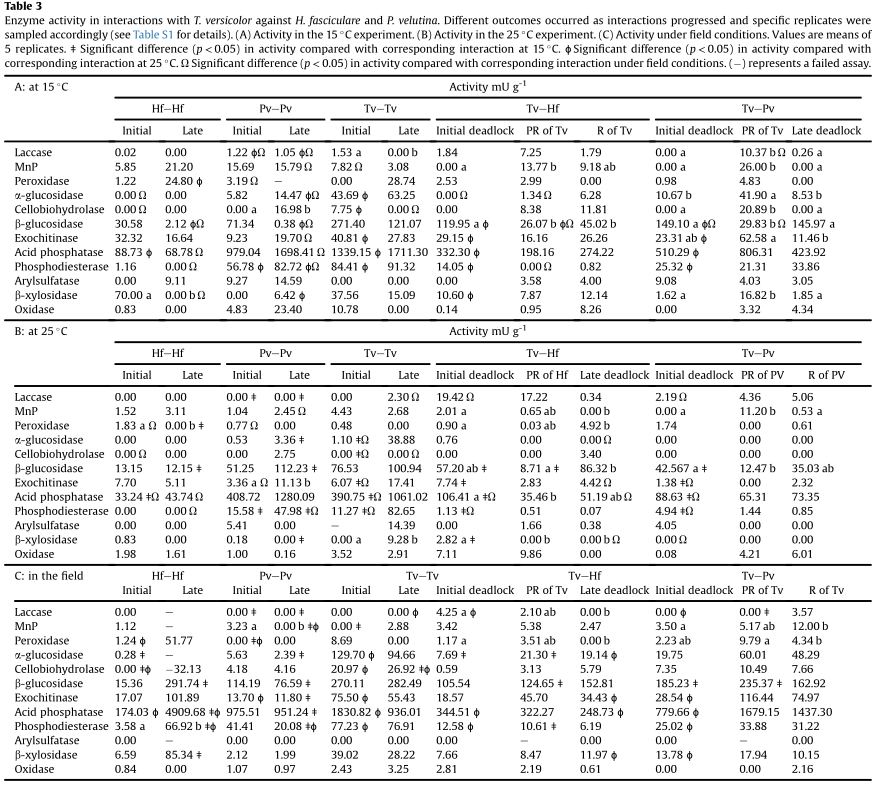
Fig. 1. Experimental set up of interactions. (A) The way in which wood blocks were paired and harvested. (B) Regions from which enzymes were extracted in interactions of T. versicolor with H. fasciculare and P. velutina (See Table S1 for specific details on sampled regions).
Three different microclimate conditions: 15℃/25℃/field mixed deciduous (predominantly beech) woodland
Harvested: 1, 14, 28, 56, and 84 d
VOCs: measured at 15 ℃ and 25 ℃ 1, 14, and 28 d (3 replicates)
Enzyme: Specific interactions and regions of interactions were chosen for enzyme analysis based on the interaction outcomes determined after reisolation onto agar.
(1) specific regions of blocks from interactions of T. versicolor with H. fasciculare and P. velutina chosen because interaction outcomes changed between different environmental conditions.
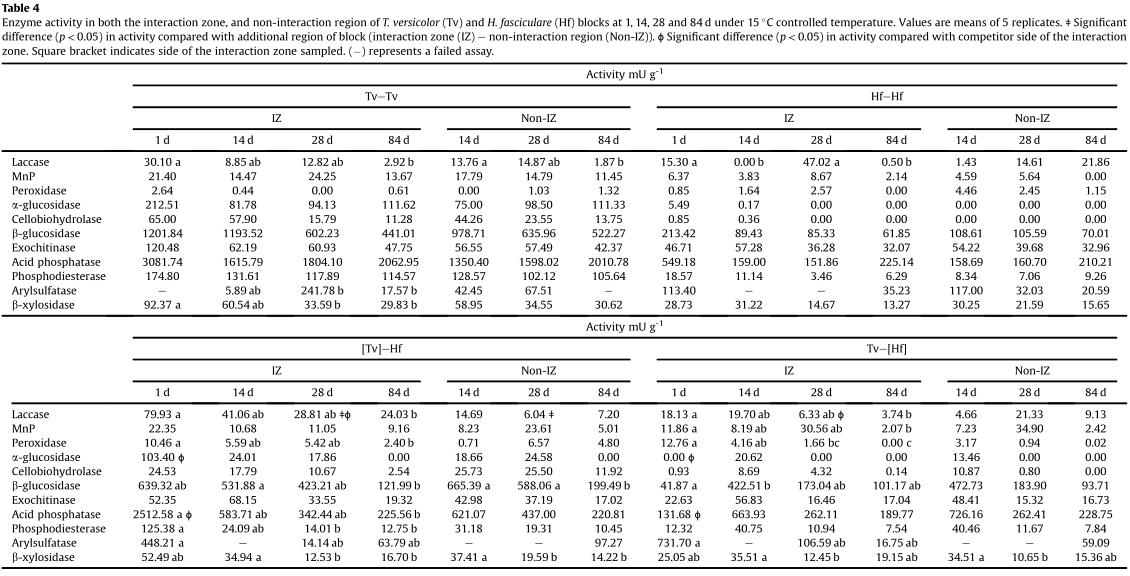
(2) all regions of interacting T. versicolor and H. fasciculare blocks over 84 d
Results:
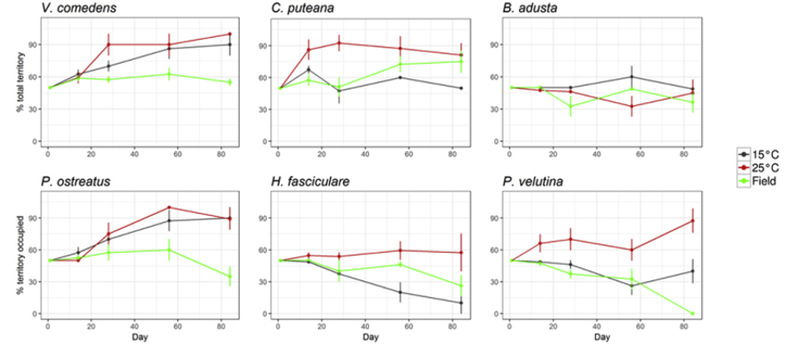
Fig. 2. Percentage territory of resource occupied by T. versicolor throughout the duration of the interaction period (84 d). Species names indicate interaction competitor.

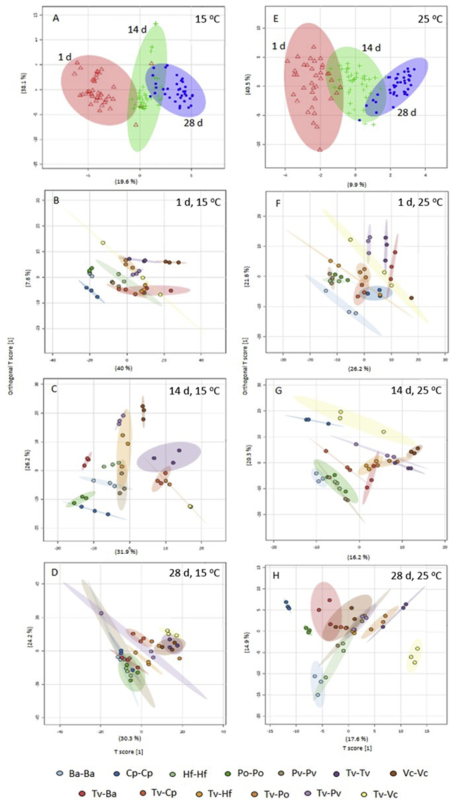
Fig. 3. OPLS-DA scores plots derived from the GC-MS spectra of interactions at 15℃(A-D) and 25℃(E-H). (A,E) Samples from all three time classes, individual pairings (B,F) 1 d after interaction set up, (C,G) after 14 d, and (D,H) after 28 d. Points represent individual samples (three independent biological replicates per pairing), and 95% confidence intervals of the means of sample groups are fitted onto the spatial ordination. Ba: B. adusta, Cp: C. puteana, Hf: H. fasciculare, Po: P. ostreatus, Pv: P. velutina, Tv: T. versicolor, Vc: V. comedens
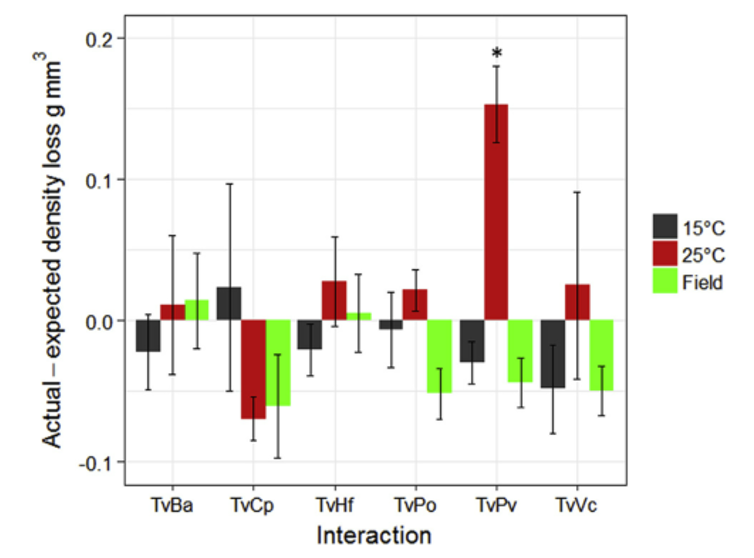
Fig. 4. Changes in block density (g mm3) following interactions (n = 5). The bars represent replicate mean ± 95% confidence interval. * indicates actual density loss minus estimated density loss (based on the rate of decay of self-pairings) was significantly (one-way t-test: p < 0.05) different to 0.
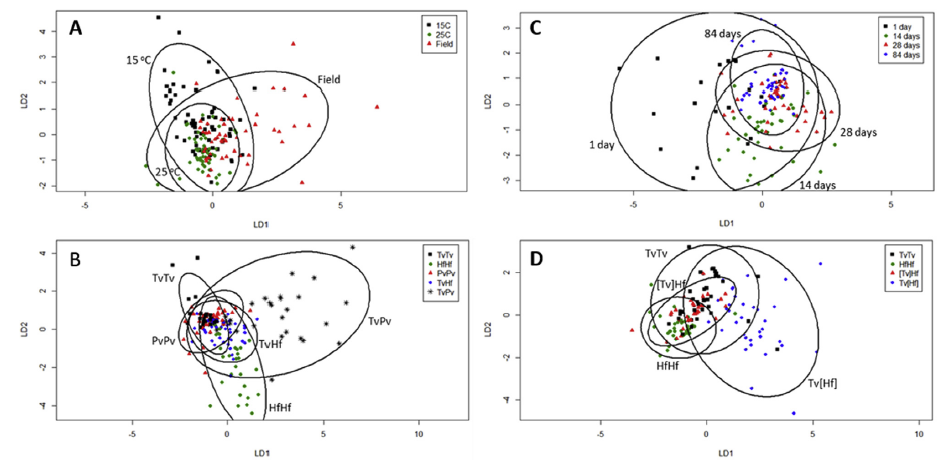
Fig. 5. Canonical analysis of principal coordinates (CAP) plots using the average activity (n ¼ 5) of enzyme profiles. CAP models were produced for (A) abiotic condition and (B) interaction combination for interactions whose outcomes differed between 15℃, 25℃and field site conditions, and for (C) sampling time and (D) region of interactions assayed for the interaction between T. versicolor and H. fasiculare over 84 d (where square brackets indicate side of the interaction zone). Ellipses show 95% confidence interval and the first two linear discriminants (LD) are shown. Tv, T. versicolor; Hf, H. fasciculare; Pv, P. velutina.
Conclusion:
It has been reported previously that temperature causes interaction outcomes to change (Hiscox et al., 2016) and that competition changes the bouquet of VOCs (Hynes et al., 2007; Evans et al., 2008; El Ariebi et al., 2016), but the present study is novel in its finding that metabolism is linked to changes in outcomes between wood decay fungi. Furthermore, tracking terminal hydrolase activity throughout the interaction between T. versicolor and H. fasciculare, revealed that its activitywas greatest during deadlock and then returned to pre-interaction levels when H. fasciculare started the process of replacement of T. versicolor and territory occupation changed. While differences in enzyme and VOC profiles between interactions with different outcomes may be incidental, fewer compounds were produced when temperature was higher suggesting that they are directly affected by temperature. Since both enzymes and VOCs contribute to diverging combative abilities and outcomes through antimicrobial activities, the relationship between metabolite production and interaction outcome under different environmental regimes is most likely explained as: temperature directly affects the production of enzymes and VOCs, and the change in metabolite production mediates interaction outcomes. This scenario explains the vital role of fungal metabolism to forest ecosystem dynamics under environmental change, and supports the first study hypothesis that the link between metabolism and interaction outcome is affected by environmental conditions. It should be mentioned, however, that enzyme activity sometimes differed between identical pairings whose interaction outcome was the same under different environmental conditions, suggesting that interaction outcomes may not be the sole cause or effect of diverging metabolic profiles.
Contact: Chen Yunru
E-mail: cherry960502@163.com