https://doi.org/10.1016/j.soilbio.2018.04.016
Highlights:
Microarthropods changed thecomposition of rhizospheric fungal communities.
A functionally diversespectrum of fungal species were stimulated.
The proportions offunctional guilds were unaffected by the microarthropods.
Introduction:
The overwhelming majority of soil fungi rely onplant-derived substances as food sources, and vegetation therefore constitutesa key factor in the bottom-up control of fungal communities. The presence and identity of living plants isespecially important for communities of those fungal groups that form part ofthe root energy channel. Another important biotic interaction of soil fungi iswith those soil-dwelling fauna that are specialized in the grazing of fungalmycelia - most prominently fungivorous nematodes, the microarthropods oribatidsand collembolans, or larger arthropods such as isopods.Depending on their bodysize and density, they may exert significant feeding pressure and suppressfungal growth or, at low intensity of feeding, induce compensatory mycelialgrowth.Thus, fungivorous soil fauna potentially regulate the composition offungal communities both at the level of functional groups and species. However,top-down control is traditionally assumed to play a negligible role in fungalassembly as compared to bottom-up factors.Recently, Crowther et al. (2013)provided direct evidence for a classical top-down effect of isopod grazing on adominant fungal taxon, which increased soil fungal diversity in microcosmalconditions. They may accelerate fungal succession on litter, regulate thespread of fungal diseases, intensify competition among functional groups ofsoil fungi or modify the composition of litter-colonizing communities of EcMF. However,surprisingly little is known about the effects of grazers in complex systemscolonized by a broad spectrum of functionally different fungal species.
The main aims of our study were therefore todetermine the effects of microarthropod presence on functionally diverse fungalcommunities in the rhizosphere. More specifically, we compared the effects amongfunctional guilds of fungi and with those of a major bottom-up factor shapingrhizospheric fungal communities – the plant's mycorrhizal host type. We assumedthat the host plant treatment would importantly affect the communities of thecorresponding mycorrhizal fungi, while the responses of the other guilds wouldbe smaller. Within this frame, we expected to find distinct levels of responseto the microarthropod presence among functionally different fungal groups.
Material and methods:
The cultivation substrate originated from a largespoil bank of a brown-coal strip mine (Sokolov region, NW Bohemia, CzechRepublic) composed of deposits of tertiary clays.The original site of thesubstrate is dominated by S. caprea with sparse understory of predominantly AMgrasses and herbs; the clay substrate is covered by a thick S. caprea litterlayer, Oe fermentation horizon of 0.5–8 cm and a thin A horizon of 0.5 cm.
At the collection of the substrate, the litterlayer was removed from the soil surface and the Oe fermentation horizon wascollected separately from the predominantly mineral material underneath, whichwas excavated to the depth of about 30 cm, and each of the two substratecomponents was homogenized. Macro- and mesofauna was eliminated from thesubstrate components by two freezing/thawing cycles of 24 h at - 40 °C, aprocedure that largely preserves microfauna and microorganisms. Defaunation byfreezing produces some side effects on the physico-chemical and microbialproperties of soils and consequently, shifts in fungal community compositioncannot be excluded. However, the microbial properties of the treated soilstended to recover within 20 days in previous studies. The mineral substrate wasfilled into round plastic pots (700 ml volume), each equipped with atransparent plastic barrier heightening the pot's margin by 30 cm to keepmicroarthropods inside the pots. The fermentation horizon was placed as a layerof 1 cm on top of the mineral substrate in order to simulate, in a smallerscale, the soil layering at the site of soil origin. Microarthropods wereextracted from the fermentation layer, collected at the same site as thecultivation substrate, with a Berlese funnel apparatus with water as preservativefluid. On average, 80 g (dry weight) of fermentation layer was used for eachfunnel set up. The mounted funnels were emptied after 24 h and the extractedmicroarthropods were added to 15 of the 30 pots. Three random extractions wereused to estimate the average number of individuals of the two mainmicroarthropod groups, collembolans and mites, added per pot, which were 90 and160, respectively. These numbers correspond to average 8000 collembolans and14,000 mites m−2, which is in the range of microarthropod densityat the site of soil origin, taking into account the lower thickness of thefermentation layer in the experiment than at the field site.
Seeds of S. caprea were germinated in autoclavedsand for two month and transferred after germination to a seedling tray with 25-ml-cellsfilled with a 1:1 (v:v) mixture of autoclaved zeolite and non-sterile spoilbank substrate from the same site as the experimental substrate. After twoweeks of greenhouse cultivation, seedlings of homogenous size were transplantedinto the experimental pots. Seeds of T. inodorum were pre-germinated andsubsequently cultivated in the same substrates as the S. caprea seedlings for10 days before transplantation into the experiment simultaneously with the S.caprea seedlings. The experiment was cultivated for 22 weeks (December to May)in a tempered greenhouse with temperature ranging between 10 °C (lowest nighttemperature) and 23 °C (highest day temperature towards the end of thecultivation period). Soil temperature was monitored and did not exceed 20 °C.The pots were watered according to the plants' needs, holes at the bottom ofthe pots, covered by fine mesh, prevented water accumulation in the pots.
The substrate was lifted en bloc from the pots andradially quartered to obtain two quarters with a plant and two without. The twoquarters without plant were divided in halves by two further radial cuts.Substrate samples were collected from all the cut surfaces, the subsamples werecarefully mixed and homogenized into one sample per pot of about 5 g freshweight and frozen. Two of the eighth parts of the substrate without plant,radially opposite, were used for the determination of microarthropod numbers.The other two eighths were suspended in water to determine the fresh weight ofroots (RFW) present in the substrate, which were picked from the suspensionwith tweezers and separated according to plant species in the pots containingboth plant species. The two quarters of the substrate containing the plantswere also suspended in water to extract the root systems and in-growing rootsof the second plant species if present. Their RFW was determined, andsubsequently, about 200–700 mg fresh roots were sampled from the intact rootsystems for the determination of root colonization and stored in 80% ethanol.
The RFW per pot was calculated by multiplying theRFW from the two substrate eighths without plant by two (to account for theroots present in the two eighths used for the determination of microacrthropodnumbers) and adding them to the RFW obtained in the two substrate quarters withplants. In the pots with two plant species, RFW was calculated for each plantspecies separately. In the pots planted with two individuals of one species,RFW per plant was calculated by dividing the total RFW per pot by two. Root dryweight (RDW) per plant was determined by multiplying the RFW per plant by aRDW-to-RFW ratio determined on one root subsample per pot. The RDW of rootsubsamples and shoot dry weights were determined after drying at 60 °C toconstant weight.
Microarthropod numbers were determined at the end ofthe experiment from the upper 5 cm of one quarter of the pot surface obtainedas described above. The microarthropods survived in most pots though in lowernumbers than added at the start of the experiment (Supplementary Table S1).This indicates that they were not prospering in the systems, possibly due tothe simplification of the original soil environment and the lower organic layerthan in field conditions. In the control treatments without microarthropodaddition, they were also occasionally recorded, but with distinctly lowerfrequency and abundance than in the treatments with microarthropod addition(Table S1).

The occurrence of single microarthropod specimensin the control treatment is attributable to the defaunation efficiency of thefreezing procedure, which is high, but usually leaves single survivingspecimens.
Root colonization was determined after Trypan bluestaining of the roots according to on 30 root segments per sample, each ca 1 cmlong, mounted on microscopic slides.The evaluations were performed under acompound microscope (Olympus BX60) at 200 × magnification.
Results:
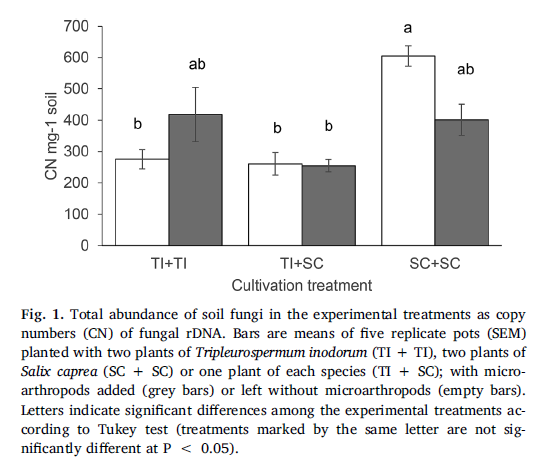
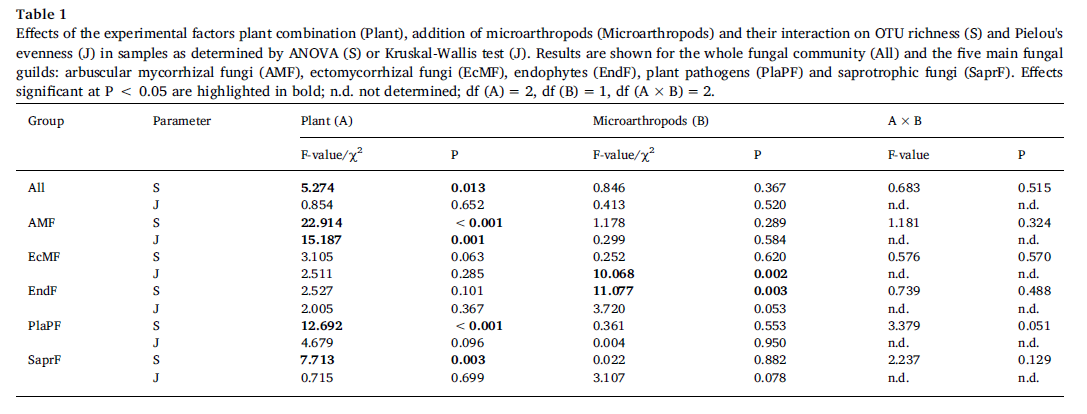
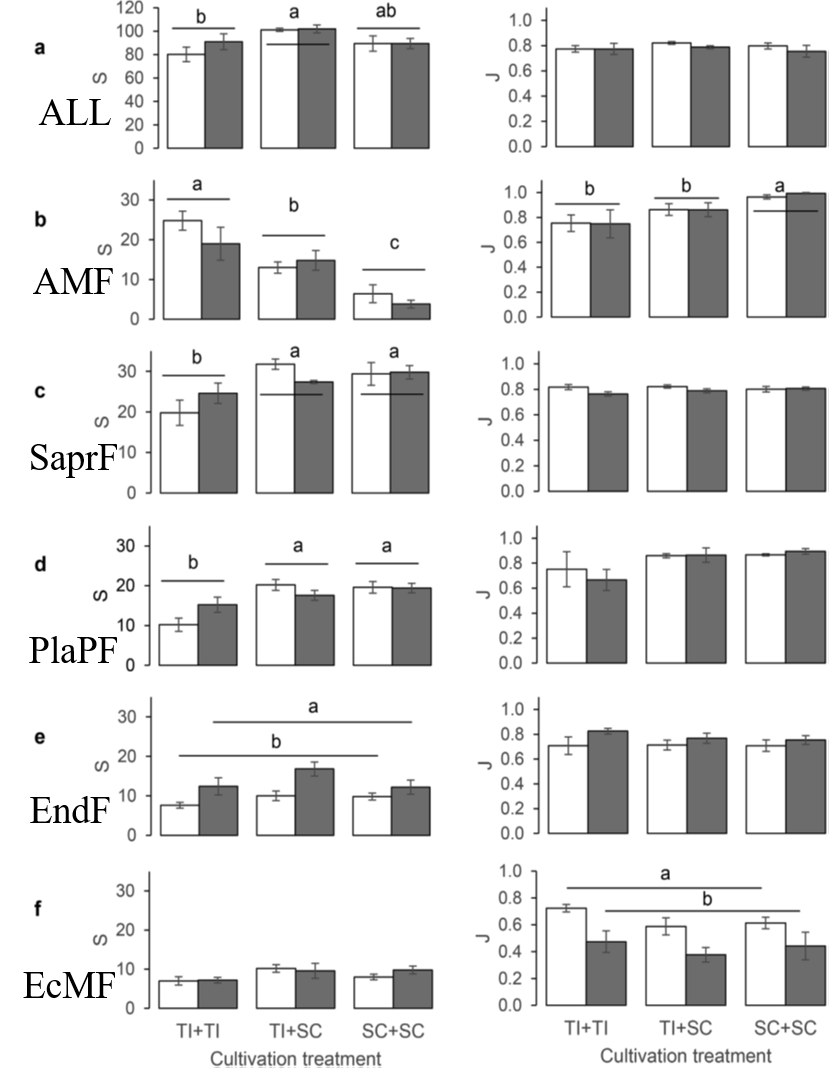
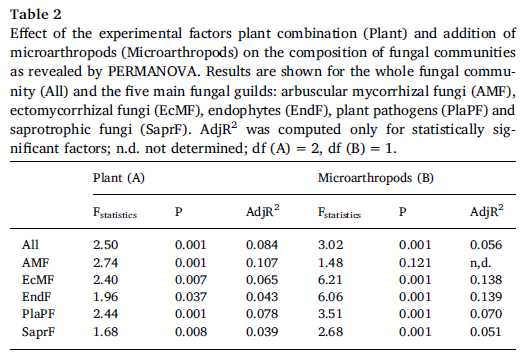
The AMFcommunity in the soil of the S. caprea monoculture had significantly lower OTUrichness and higher evenness than communities established in the presence of T.inodorum (Fig. 2b). Plant combination also significantly affected the communitycomposition of AMF (Table 2), while microarthropods had no significant effectat all on the AMF communities. Similarly as in AMF, the OTU richness of SaprFand PlaPF communities was affected by plant combination only (Table 1), but thespecific differences between treatments were opposite as in AMF: OTU richnesswas significantly lower in the T. inodorum monoculture than in the presence ofS. caprea (Fig. 2c and d). In contrast to the previously described groups, thecommunities of EcMF and EndF were more affected by microarthropod presence thanby plant combination: Microarthropods increased the OTU richness of EndF (Fig.2e) and decreased the community evenness of EcMF (Fig. 2f), while plantcombination had no significant effect on the diversity parameters of eitherguild (Table 1). The community composition of the EcMF and EndF guilds wassignificantly affected by both experimental factors, with microarthropods explainingdistinctly higher proportion of total variance than cultivation system (Table2).
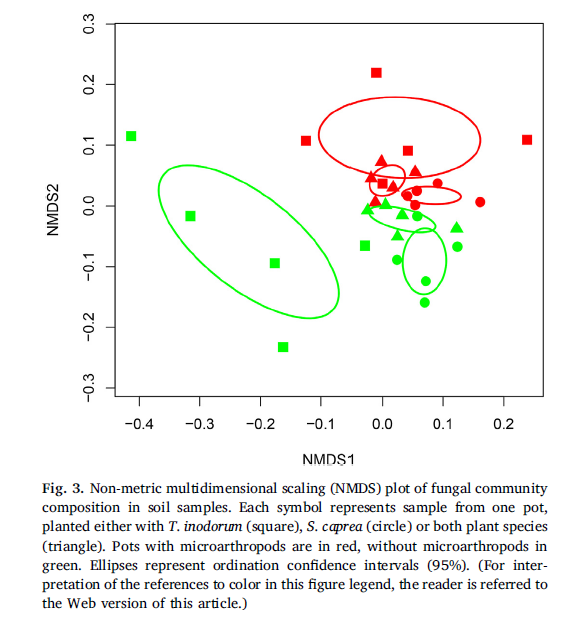
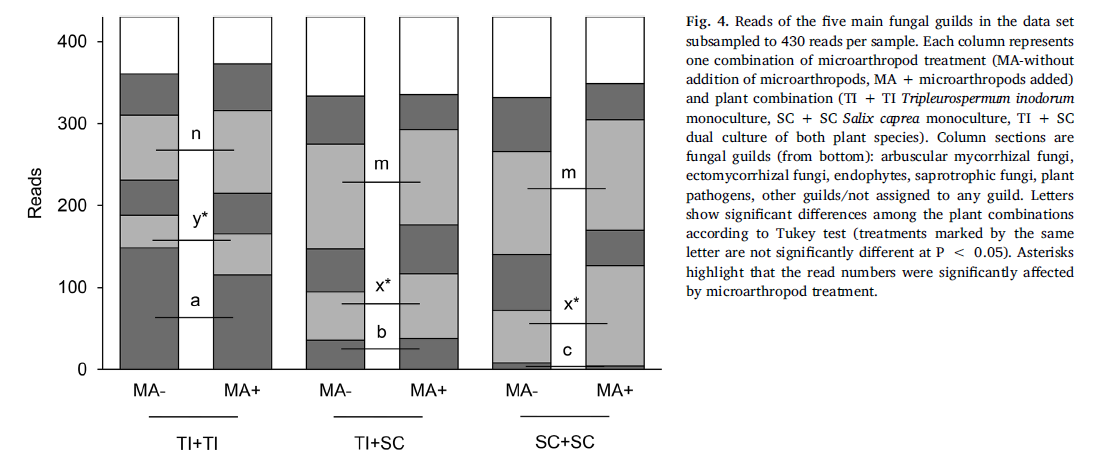
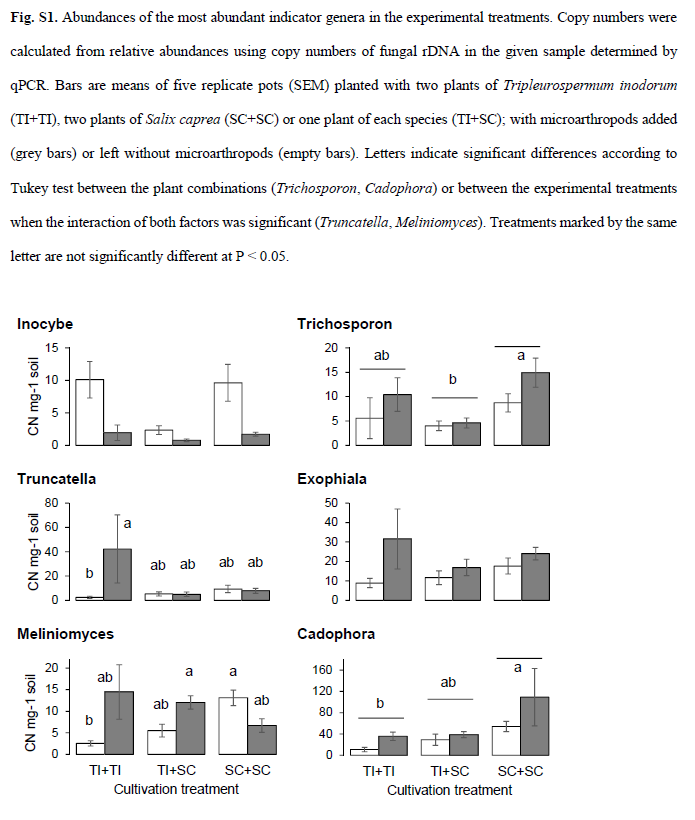
As expected,the differences were driven by AMF - significant indicator guild for the T.inodorum monoculture, and EcMF - significant indicator guild for the S. capreamonoculture (cluster indicator values 0.5525 and 0.3942, respectively,P = 0.001 and 0.009). Nevertheless, the proportion of EcMF in the fungalcommunity was significantly higher in the presence of microarthropods (Fig. 4).Letters show significant differences among the plant combinations according toTukey test (treatments marked by the same letter are not significantlydifferent at P < 0.05). Asterisks highlight that the read numbers weresignificantly affected by microarthropod treatment.
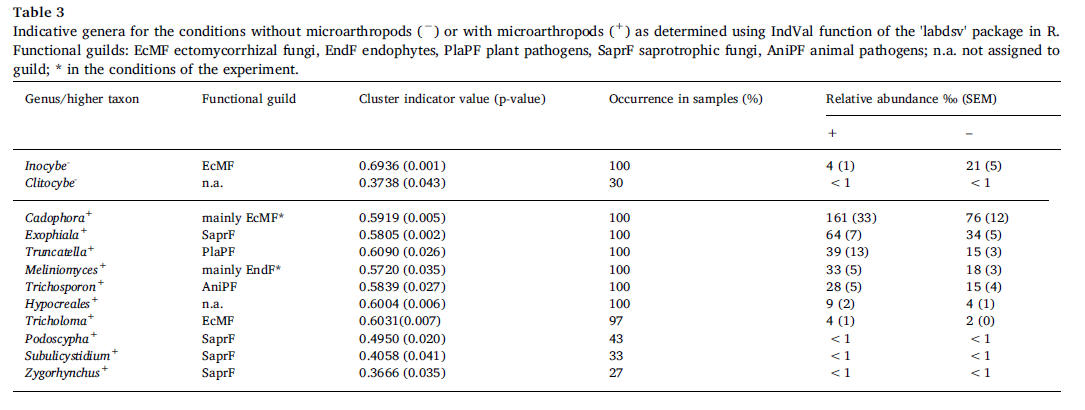
Two fungalgenera were identified as indicative for the conditions withoutmicroarthropods, nine genera and one higher order taxon for the conditions withmicroarthropods (Table 3).The most abundant genus in the whole data set, in ourconditions predominantly EcM Cadophora, was found indicative for the conditionswith microarthropods, increasing its average relative abundance from 8% to 16%.Another EcM genus, Inocybe, decreased its relative abundance in the presence ofmicroarthropods from 2.1% to 0.4%. The other genera with average relative abundancehigher than 1% were all more abundant in the presence of microarthropods:Exophiala (SaprF), Truncatella (PlaPF), Meliniomyces (predominantly EndF in ourconditions) and Trichosporon (animal pathogen) (see Table 3 for details).
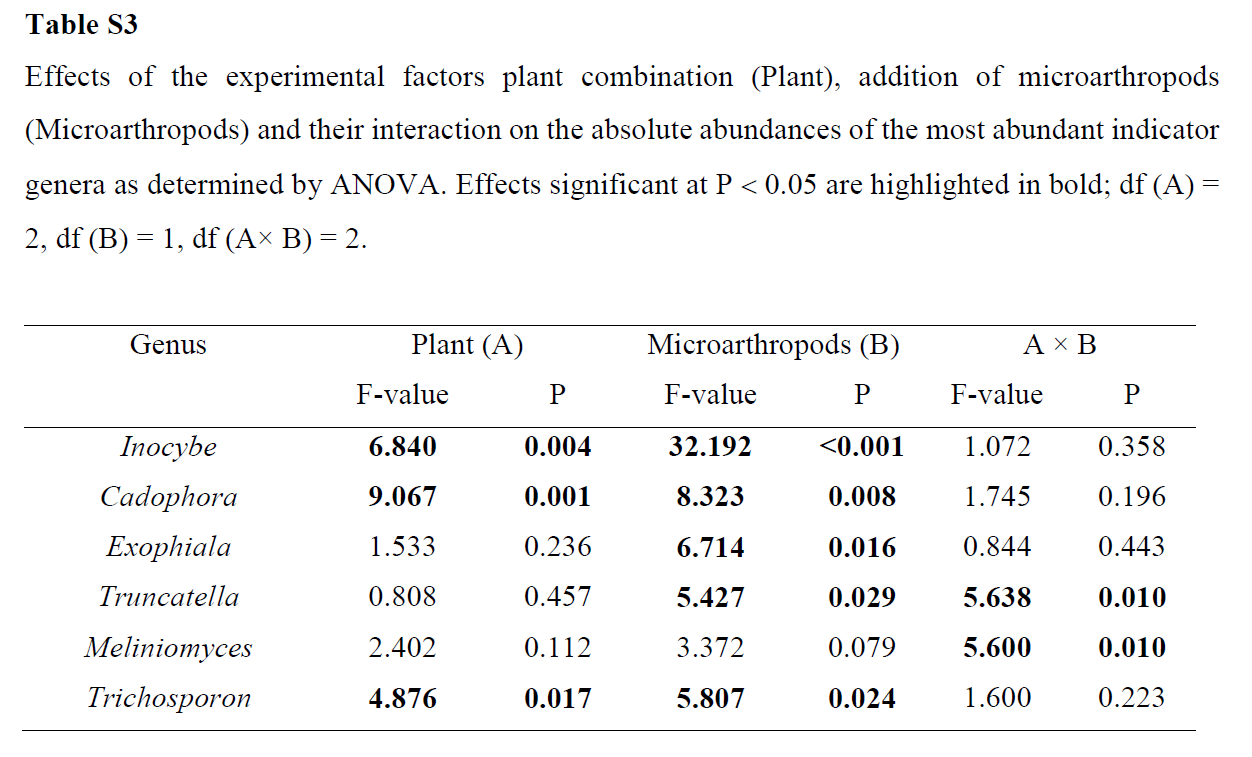
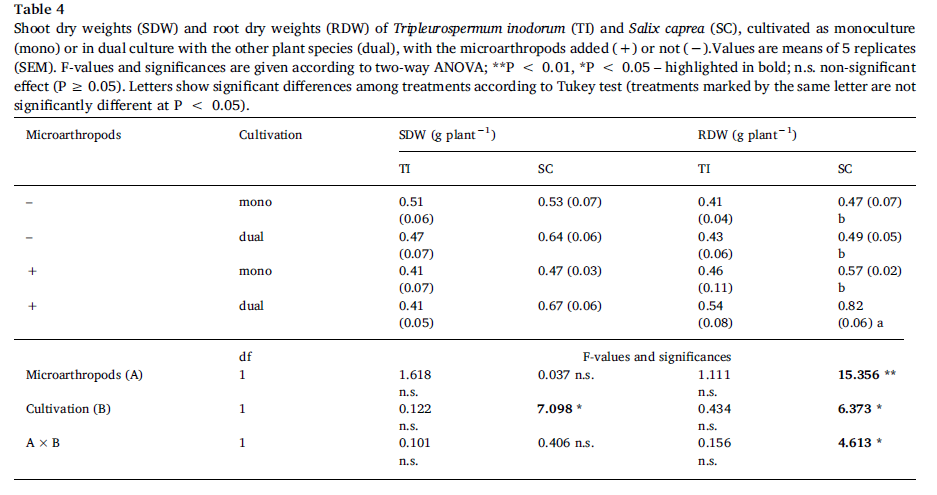
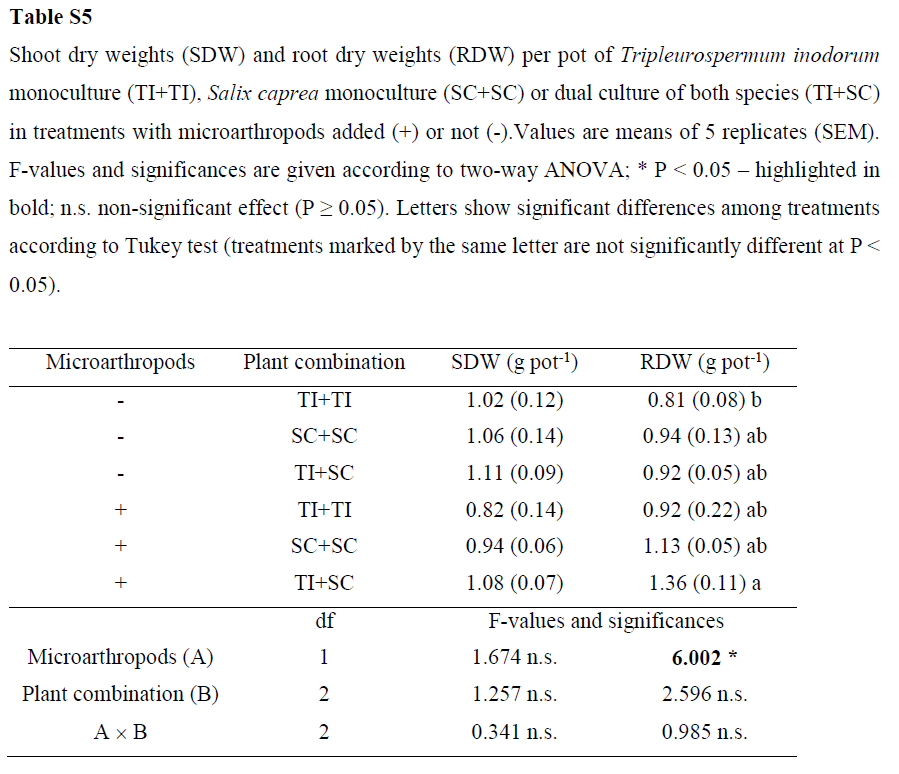
The per-plantshoot biomass of S. caprea was significantly higher in dual cultivation with T.inodorum than in S. caprea monoculture (Table 4).The root biomass of S. capreawas also significantly higher in the presence of microarthropods, and thedifference was more pronounced in dual cultivation than in monoculture. Theper-plant biomass of T. inodorum was unaffected by plant combination ormicroarthropods. The total shoot biomass per pot was also unaffected by theexperimental factors, while the total root biomass per pot was significantlyhigher in the presence of microarthropods (Supplementary Table S5).
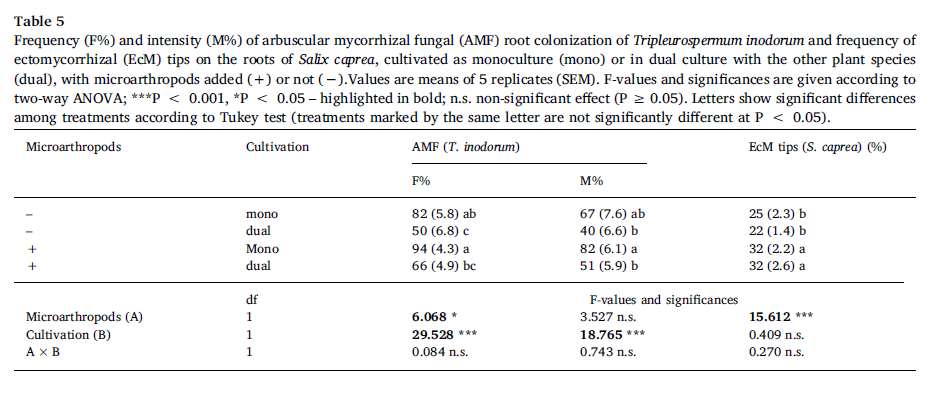
F% and M% of AMfungal colonization in T. inodorum roots were significantly higher in T.inodorum monoculture than in dual cultivation with S. caprea, the presence ofmicroathropods significantly increased F% (Table 5). Plant combination had noeffect on the frequency of EcM root tips on S. caprea roots, while the presenceof microarthropods significantly increased it (Table 5).
Discussion:
Both host plantcombination and microarthropod presence significantly affected composition ofthe fungal communities in the rhizospheres of our experiment. Only one genuswith relative abundance higher than 1% (EcM Inocybe sp.) was indicative for theconditions without microarthropods, i.e. was significantly reduced in theirpresence. In contrast, five genera of more than 1% relative abundanceapproximately doubled their relative abundances in the presence ofmicroarthropods (Table 3), and four of them also significantly increased theirabsolute abundances. The absence of concomitant reduction of a dominant taxonsuggests that this response was not related to competitive release asdocumented previously, but rather to relative stimulation by the faunapresence. One possible explanation for these findings is the differentialability of fungal taxa to regrow from fragmented mycelia, as microarthropodscause mechanical stress and fragmentation of fungal hyphae by grazing on themycelia and soil perturbation. Fragmented mycelia are also used as inocula forfungal cultivation, which suggests that culturability of fungal speciesindicates higher tolerance to fragmentation of fungal hyphae. In our study,species of the genus Inocybe, which are not culturable in vitro, respondednegatively to microarthropod presence, while the taxa that increased theirrelative abundances in the presence of microarthropods were repeatedly isolatedfrom soil or host tissue into pure cultures. Furthermore, fungal speciestolerant to mycelial fragmentation are more likely to be dispersed within thesoil by microarthropods. Generally, microarthropods can promote the dispersionof a broad range of fungi to new microsites within soil. However, mechanicalstress caused by microarthropods may also have played a role in the response ofthis species, as it should favour yeasts with unicellular lifestyle, such asTrichosporon sp., as compared to filamentous fungi. Finally, we may attributethe responses of some taxa to being grazed by the microarthropods. This mayparticularly apply to Exophiala sp. and Cadophora sp. that produce darkpigmented mycelia, preferentially consumed by fungivorous microarthropods.
Relativestimulation of the Cadophora and Meliniomyces genera belonging to Rhizoscyphusericae aggregate is noteworthy because it substantially contributed to the highproportion of variation explained by microarthropod presence within the EcMFand EndF guilds. The high responsiveness of this taxonomic unit, which cannotbe unambiguously assigned to one fungal guild, corroborates the assumption thatthe microarthropod effects were driven by responses of specific fungal taxaindependently of their trophic position or functional characteristics. Theassignment of this taxonomic group into two functional guilds also illustrateshow the effect of faunal grazers on fungal diversity depends on the dominancestatus of the most responsive taxa. Within EndF, microarthropods stimulatedseveral OTUs of low relative abundance (assigned to Meliniomyces sp.) and thusincreased the OTU richness per sample and tended to increase the communityevenness (Table 1, Fig. 2f). Within EcMF, in contrast, microarthropodsstimulated the OTU with the overall highest relative abundance (Cadophorafinlandica) and consequently, decreased the evenness of the EcMF community.
As mentionedabove, microarthropods did not induce major functional shifts in the fungalcommunity as they had no significant effect on its composition by functionalguilds. However, the significant increase of root colonization in both guildsof mycorrhizal fungi (AMF and EcMF) suggests that microarthropods significantlystimulated their activity. This is consistent with previous reports on positiveeffects of soil fauna on the functioning of AM and the reasoning of Gange(2000) that this should be partly attributed to stimulation of hyphal growth.
In conclusion,our results point to the importance of targeting complex fungal communities toobtain a correct picture of the impact of fungal grazers. Relatively largeeffects within EcMF and EndF were counterbalanced by smaller effects in theother fungal guilds, and microarthropods did not significantly shift the ratiosof the fungal guilds within the communities. This indicates that the functionalcomposition of rhizospheric fungal communities is mainly governed by bottom-upfactors related to plant identity and the effect of microarthropods isfunctionally neutral. However, we also show that microarthropods cansignificantly change the taxonomic composition of rhizospheric fungalcommunities by increasing the relative abundance of some taxa. We hypothesisethat this effect is attributable to differential responses of fungal taxa tomechanical disturbance, dispersion within the soil or low-intensity grazing bythe microarthropods. We advocate that studies as the one presented here provideimportant indication on which fungal taxa and traits to concentrate detailedinvestigations of the soil fauna-fungi interactions in microcosmal conditions.
chenyunru