Resolving fungal and bacterial groups within the microbialdecomposer community is thought to capture disparate life strategies for soilmicrobial decomposers, associating bacteria with an r-selected strategy forcarbon (C) and nutrient use, and fungi with a K-selected strategy. Additionally,food-web model-based work has established a widely held belief that thebacterial decomposer pathway in soil supports high turnover rates of easilyavailable substrates, while the slower fungal-dominated decomposition pathway supportsthe decomposition of more complex organic material, thus characterizing the biogeochemistryof the ecosystem.
Article 1:
Title: Revisiting the hypothesisthat fungal-to-bacterial dominance characterizes turnover of soil organicmatter and nutrients
Downloadwebsite:https://esajournals.onlinelibrary.wiley.com/doi/full/10.1890/14-1796.1
Maincontents:
Author used a field experiment, the Detritus Input and RemovalTreatments, or DIRT, experiment (Harvard Forest Long-Term Ecological ResearchSite, USA) where litter and root inputs (control, no litter, double litter, orno tree roots) have been experimentally manipulated during 23 years, generatingdifferences in soil C quality. We hypothesized (1) that δ13Cenrichment would decrease with higher soil C quality and that a higher Cquality would favor bacterial decomposers, (2) that the C mineralized infungal-dominated treatments would be of lower quality and also depleted in δ13C relative to bacterial-dominatedhigh-quality soil C treatments, and (3) that higher C mineralization along withhigher gross N mineralization rates would occur in bacterialdominated treatmentscompared with more fungal-dominated treatments. The DIRT treatments resulted ina gradient of soil C quality, as shown by up to 4.5-fold differences betweenthe respiration per soil C between treatments. High-quality C benefited fungal dominance.Further, there was no difference between the δ13C O2 produced by afungal compared with a bacterial-dominated decomposer community. There weredifferences in C and N mineralization between DIRT treatments, but these werenot related to the relative dominance of fungal and bacterial decomposers.
1. Fungaland bacterial growth.There were small but systematic effects oftreatments and horizon, with an interaction between the factors for fungalgrowth per unit SOC (Fig. 1a, b). Higher litter input appeared to stimulatefungal growth; however, a clear effect was the high rate of fungal growth inthe OA treatment, with fungal growth rates being higher overall in the Ocompared with the M horizon. The bacterial growth per unit SOC was affected bytreatment but not by horizon (Fig. 1c, d). This yielded a ratio between fungal andbacterial growth that was affected by treatment but not horizon (Fig. 1e, f).Again, we observed a trend where treatments with litter inputs (2X, Co, OA, T)were higher than those without litter input (0X, T0X) suggesting a correlationbetween fungal dominance and litter input.
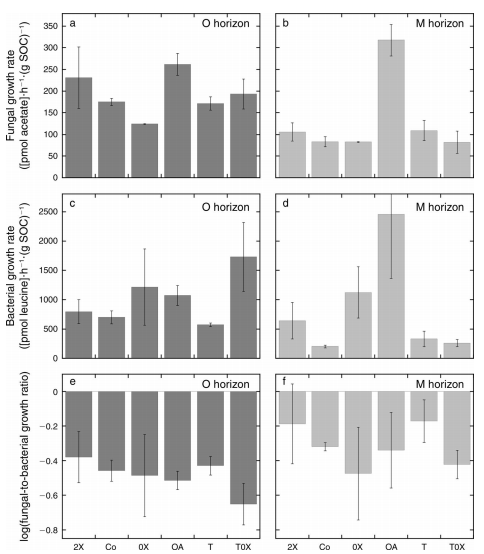
Fig.1 (a, b) The fungal growth rate as determined usingthe acetate incorporation into ergosterol method, (c, d) the bacterial growthrate as determined using the leucine incorporation into extracted bacteriamethod, and (e, f) the logarithm of the ratio between fungal-to-bacterialgrowth. The O (a, c, and e) and M (b, d, and f) horizon results are presentedseparately.
2.Respiration and gross N transformation. The respiration rate per unit SOC was affected by bothtreatment and horizon (Fig. 2a, b). The NH4+production per unit SOC estimates gave a very similar, and yet clearer picture(Fig. 2c, d). Rates were higher in the O than M horizons, and treatments withlitter input (2C, Co, OA, and T) were all clearly higher than those withoutlitter input (0X and T0X) in the O horizon, with no obvious treatmentdifferences in the M horizon. No significant treatment or horizon differenceswere observed for NH4+ consumption per unit SOC estimates (Fig. 2e,f).
3. Microbial biomass. The microbial biomass per unit SOC, asmeasured by SIR, depended on treatment but only marginally on soil horizon. TheOA treatment stood out with an approximately twofold higher SIR biomass thanthe other treatments, which were not distinguishable. The OA treatment stoodout with an approximately twofold higher SIR biomass than the other treatments,which were not distinguishable. The bacterial PLFA concentration per unit SOCwas unaffected by treatment and horizon. The fungal PLFA concentration per unitSOC depended on both treatment and horizon, with an interaction showing that treatmenteffects were larger in the O horizon.
4. Microbial PLFA composition and δ13C content. The δ13Cenrichment of PLFA markers did not depend on treatment or horizon (Fig. 5).There were systematic differences between the enrichment of different PLFAmarkers (Fig. 5), where the δ13C enrichment ranged betweenrelatively enriched markers ,When aggregated into the taxonomic categorieslisted in Fig. 5, there were significant differences between groups with d13Cenrichment ranging from highest to lowest as G-/AMF, Actin, G+, G-/F, F and G- (taxonomicgroups defined in Fig. 5).
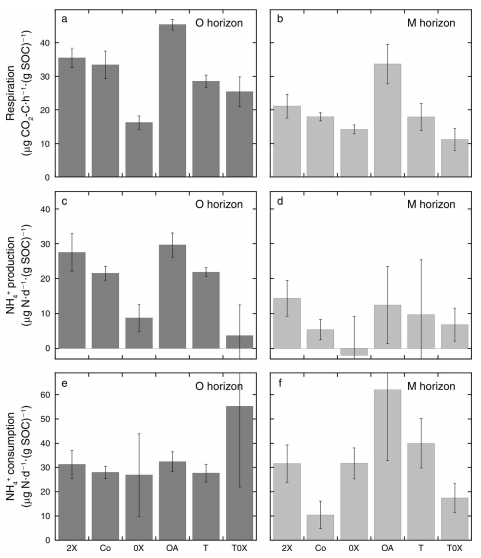
Fig.2The microbial rates of (a and b) C and (c–f) N, andconsumption, panels e and f transformation. The O (panels a, c, and e) and M(panels b, d, and f) results are presented separately. Values represent themean 6 SE of six (Co) or three (other treatments) field replicates, and arepresented per soil organic carbon (SOC).
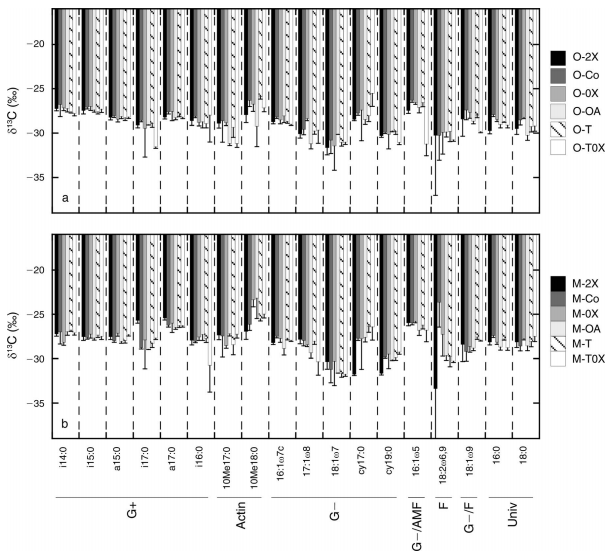
Fig.3The δ13C in extracted PLFA markers in the (a)O and (b) M horizons. The treatments are color coded according to legend.Markers associated with specific microbial groups are noted as G+ (grampositive bacteria), G- (gram negative bacteria), Actin (actinomycetes), AMF(arbuscular mycorrhizal fungi), F (saprotrophic fungi), and Univ (universal).
Thestudy find no support for the hypothesized differences between detritalfood webs dominated by bacteria compared to those dominated by fungi. Rather,an association between high-quality soil C and fungi emerges from our results.Consequently there is a need to revise our basic understanding for microbialcommunities and the processes they regulate in soil.
Article 2:
Title: Linking bacterial andeukaryotic microbiota to litter chemistry: Combining next generation sequencingwith 13C CPMAS NMR spectroscopy
Downloadwebsite:https://www.sciencedirect.com/science/article/pii/S0038071718303882?via%3Dihub
Maincontents:
Microbial successionover decomposing litter is controlled by biotic interactions, dispersallimitation, grazing pressure, and substrate chemical changes. Recent evidencesuggests that the changes in litter chemistry and microbiome duringdecomposition are interdependent. However, most previous studies separatelyaddressed the microbial successional dynamics or the molecular changes ofdecomposing litter. Here, we combined litter chemical characterization by 13CNMR spectroscopy with next generation sequencing to compare leaf litter chemistryand microbiome dynamics using 30 litter types, either fresh or decomposed for30 and 180 days.
1. Litter mass loss andassociated chemistry. Litter mass loss varied among litter types, and for allspecies, over decomposition time (Fig. 1). Massloss was initially very rapid for A.mollis, C. emerus, H. helix, and F. ornus, and slower for Q.ilex, F. sylvatica, and P. halepensis, showing intermediatevalues for P. nigra, R. pseudoacacia and F. drymeia. However, as decomposition was proceeding, mass lossprogressively declined for all litters, especially for A. mollis, C. emerus, andH. helix.13C NMR spectra revealedsignificant and consistent differences among litter species, as well as changesin the relative abundance of different C types during decomposition (Fig. 2).
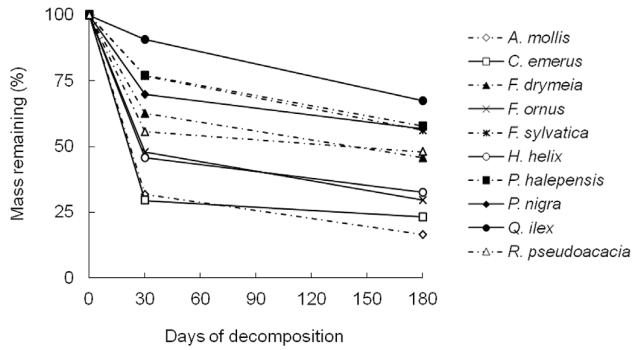
Fig. 1.Dynamics of litter mass remaining during 180 days of decomposition. Data referto mean of 10 replicates for each species.
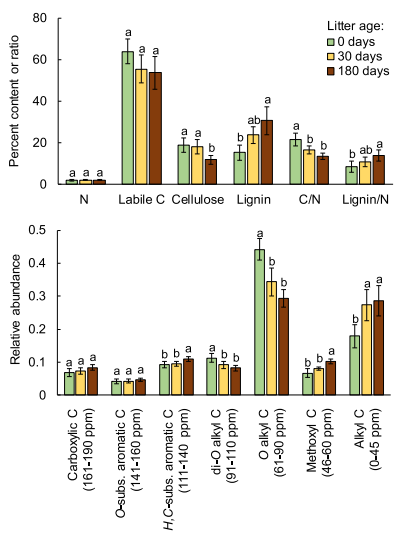
Fig. 2.Dynamics of litter nitrogen (N), labile C, cellulose and lignin percentcontent, C/N and lignin/N ratios (top), and C-types corresponding to 13CCPMAS NMR regions (bottom) during 180 days of decomposition.
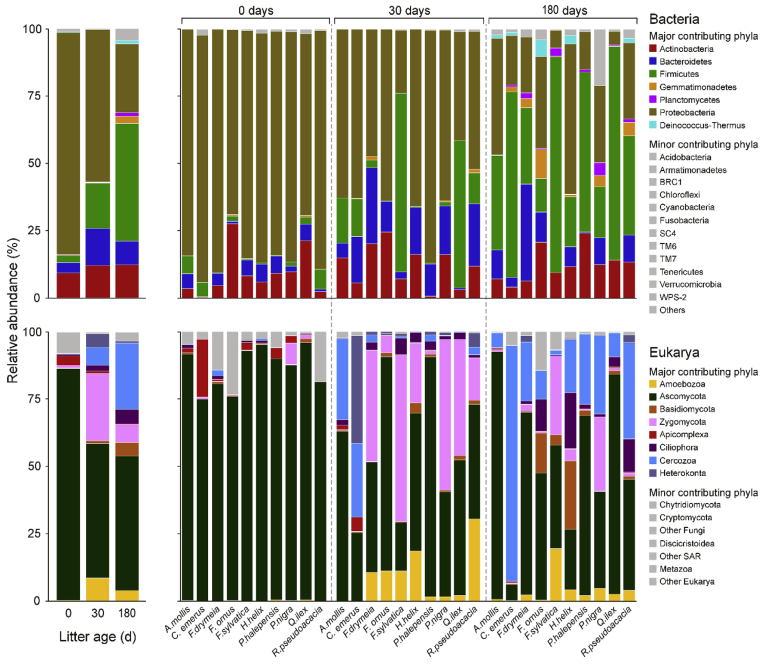
Fig. 3.Relative abundance of bacterial (top) and eukaryotic (bottom) phyla in littersamples of different ages, either averaged (left) or separately reported for 10different litter species (right).
2. Bacterial succession. An overview of bacterial succession isreported in Fig. 3. Proteobacteriadominated on undecomposed litter (mean ± standard error of relative abundancewas 83.0 ± 2.6%), followed by Actinobacteria, Bacteroidetes and Firmicutes (9.2± 2.7%, 3.9 ± 0.8% and 2.7 ± 0.8%). As decomposition was proceeding,Proteobacteria were largely substituted by Firmicutes. Actinobacteria andBacteroidetes generally increased with litter age, but the response was litterspeciesspecific.
3. As forprokaryotes, Fig. 3 shows eukaryotic successional pattern during litterdecomposition. Members of the phylum Ascomycota dominated the eukaryoticcommunity of all undecomposed litter materials (mean ± standard error ofrelative abundance 86.6 ± 2.5%).
4. Linking microbial succession to litterchemistry. Correlation networks between litter microbiota and chemical traits provideda synthetic picture of the interdependence of microbial community compositionand litter chemistry, as related to leaf type and age (Fig.4). In undecomposedmaterials, the large predominance of Proteobacteria was positively associatedwith the substrate C/N ratio, cellulose content, relative fraction of O-alkyl Cand, to a lesser extent, Alkyl C (Fig. 4A). Proteobacteria abundance was alsonegatively associated with litter N content, carboxylic and methoxyl Cfractions (Fig. 4B).
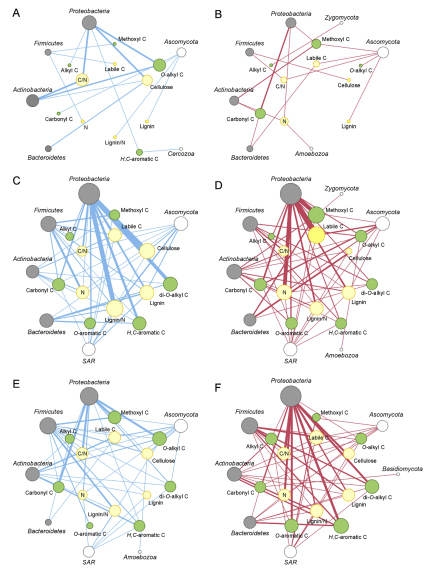
Fig. 4. Relationship between microbiome and litter chemistry for10 different litter materials either undecomposed (A–B) or after 30 (C–D) and 180(E–F) days of decomposition. In each panel, data refer to correlation networkbetween Bacteria and Eukarya phyla (dark grey and white nodes, respectively)and litter chemical traits (green and yellow nodes for 13C NMR-based C typesand elemental/proximate parameters, respectively). Node size islog-proportional to the total number of significant correlations observedwithin each phylum or litter trait. Linewidth is proportional to the number ofOTUs in a given phylum significantly associated to a specific litter trait,either positively (blue lines, A, C, F) or negatively (red lines, B, D, E).Phyla with no OTUs associated to litter chemistry or litter traits unrelated tolitter microbiome are not shown. (For interpretation of the references tocolour in this figure legend, the reader is referred to the Web version of thisarticle.)
Conclusion:
Thestudyshowed thatdecomposing litter undergoes significant chemical transformations withassociated microbiological successional shifts. Despite the wide chemical rangeof litter materials purposely selected for our decomposition experiment, mostlitter types remarkably shared similar molecular changes and successionaltrajectories. A such, our findings suggest a common pattern in the microbial communitycompositional response to the chemical changes that occur during decomposition.
Article 3:
Title:Mechanisms linking fungal conditioning of leaf litter todetritivore feeding activity
Downloadwebsite:https://www.sciencedirect.com/science/article/pii/S0038071715003806?via%3Dihub
Maincontents:
Saprotrophic fungi are one of the primary groups oforganisms responsible for decomposition and nutrient cycling processes. Asidefrom their direct effects on litter decomposition, these organisms may indirectlyaffect litter invertebrate activity via fungal-mediated conditioning (i.e.‘priming’) of litter substrates. While this priming effect is generally assumedto take place, this assumption is based on correlative observations and feedingpreference trials rather than experimental discrimination of the mechanismsdriving the process. Author used a microcosm experiment of brown-rot (Fistulina hepatica, Fomitopsis lilacinogilva) and white-rot (Pycnoporus coccineus) fungi on two sclerophyllous leaf litters (Eucalyptus loxophleba and Acacia acuminata) to test if afungal-mediated change in plant litter increased the nutritive value of thefood source, and if a change in cell-wall (structural) compounds reduced the toughnessof leaves, resulting in increased feeding activity by invertebrates.
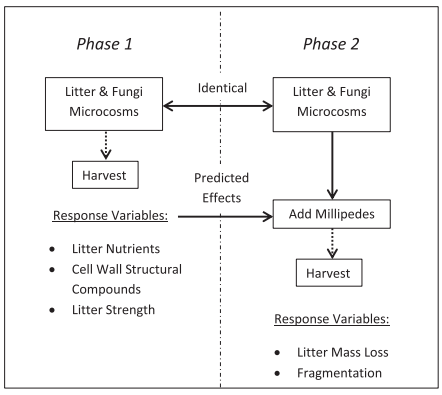
Fig. 1.The experimental protocol depicting the two-phased approach we used to separatethe effects of fungal colonization on (1) leaf chemistry and mechanical strength,and (2) millipede feeding activity.
Results:
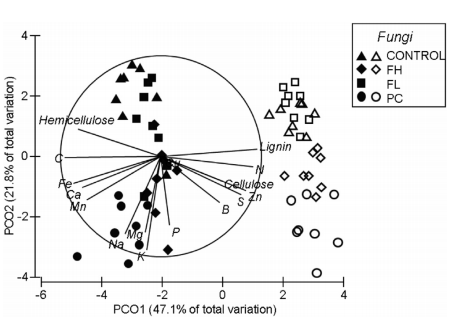
Fig. 3. Principal Coordinates Analysis (PCO) biplot showingvariation in cell wall structural compounds and nutrients among fungitreatments and leaf litter types. Biplot was constructed using normalized dataand Euclidean distance. Points represent individual microcosms; closed symbolsshow microcosms containing Eucalyptus loxophlebaleaf litter and open symbols show microcosms containing Acacia acuminate leaflitter. CONTROL represents the control treatment; FH, Fistulina hepatica; FL, Fomitopsislilacinogilva; and PC, Pycnoporuscoccineus. The Pearson correlations overlay highlights the linearrelationships between the chemical response variables and PCO axis 1 and 2; thelength and direction of the vectors indicates the strength and sign of theserelationships.
2. The primingeffect of fungi on millipede feeding activity.There were significant effects of litter species, and fungal treatments onmillipede mediated mass loss and fragmentation, but no significant interactionbetween litter species and fungi on these response variables. Millipede mortalitywas low and feeding was observed across all microcosms, although to a lesserextent than expected. Pairwise comparisons showed that the only fungustreatment in which there was significantly greater mass loss and leaf fragmentation than in the control forboth Acacia and Eucalypt microcosms was the PC treatment.
The final most parsimonious SEM modelfor PC-Eucalypt (Fig. 2a) had acceptable goodness of fit indices. There was no significant difference between the predictedcovariance structure of the final model and the observed covariance structurein the data. The final, most parsimonious SEM for PC-Acacia had acceptable goodnessof fit indices, and the predicted covariance structure of the final model and theobserved covariance structure of the data did not differ significantly.
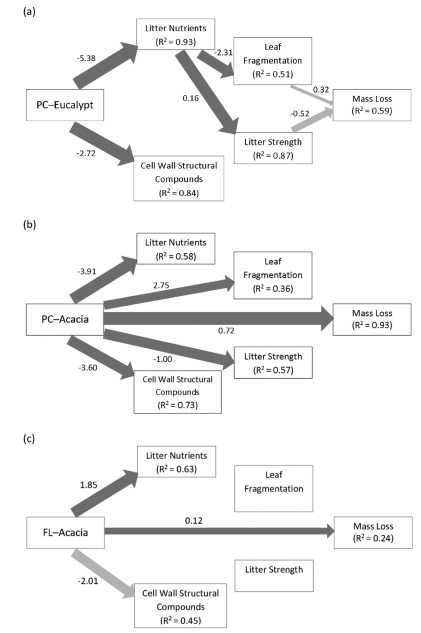
Fig. 2. The most parsimonious structural equation models for Acacia acuminata (Acacia) and Eucalyptus loxophleba (Eucalypt)discriminating direct versus indirect mechanistic pathways through which fungalcolonization and subsequent changes in leaf litter structure and leaf litterchemistry influence litter strength, and millipede mediated leaf fragmentationand mass loss. Results of model fitting for each fungi-litter species treatmentare as follows: (a) PC-Eucalypt; (b) PC-Acacia; (c) FL-Acacia. Litter structureand litter chemistry are composite variables composed of axis scores ofprinciple components analyses. The arrows represent causal paths from predictorto response variables; dark arrows indicate significant effects (P < 0.05),light arrows represent effects that were not significant, and arrow widthcorresponds directly to standardized path coefficients. Numerical valuesassociated with solid arrows represent unstandardized partial regression coefficientsand indicate whether the relationship is positive or negative. R2values associated with response variables indicate the proportion of variationexplained by the relationships with other variables.
Conclusion:
The studysuggest that, in theearly stages of decomposition, fungal priming effects on leaf litter nutrientsare a more important determinant of litter-invertebrate interactions thanchanges to litter structural compounds. Furthermore, different species of fungiinteract with different litter types and invertebrates in functionallydifferent ways.
Contact: CaoTingting
E-mail:1274467369@qq.com