https://doi.org/10.1073/pnas.1811269115
Abstract
Bacteria and fungidrive decomposition, a fundamental process in the carbon cycle, yet theimportance of microbial community composition for decomposition remainselusive. Here, we used an 18-month reciprocal transplant experiment along aclimate gradient in Southern California to disentangle the effects of themicrobial community versus the environment on decomposition. Specifically, wetested whether the decomposition response to climate change depends on themicrobial community. We inoculated microbial decomposers from each site onto acommon, irradiated leaf litter within “microbial cages” that prevent microbialexchange with the environment. We characterized fungal and bacterialcomposition and abundance over time and investigated the functional consequencesthrough litter mass loss and chemistry. After 12 months, microbial communitiesaltered both decomposition rate and litter chemistry. Further, the functionalmeasurements depended on an interaction between the community and its climatein a manner not predicted by current theory. Moreover, microbial ecologistshave traditionally considered fungi to be the primary agents of decompositionand for bacteria to play a minor role. Our results indicate that not only doesclimate change and transplantation have differential legacy effects amongbacteria and fungi, but also that bacterial communities might be lessfunctionally redundant than fungi with regards to decomposition. Thus, it maybe time to reevaluate both the role of microbial community composition in itsdecomposition response to climate and the relative roles of bacterial andfungal communities in decomposition.
Materials and Methods
Field Experiment.
The five field sites(desert, scrubland, grassland, pine–oak, and subalpine, named for the vegetationpresent) were selected to represent a wide temperature and precipitation rangewithin Southern California (Fig. 1AandSI Appendix, Table S1). On October 19, 2015, we deployed 300 litterbags containingirradiated grassland litter inoculated with one of the microbial communitiesfrom each of the five sites into each site. We selected litter from themidelevation grassland site as a common substrate, because the site isintermediate in temperature and precipitation and has been intensivelycharacterized as part of the long-term Loma Ridge Global Change Experiment. Thegrassland is dominated by the annual grass generaAvena,Bromus,andLolium; the annual forb generaErodiumandLupinus; and the nativeperennial grassNassella pulchra.
The grassland litterwas homogenized by grinding in a coffee grinder. In addition to improvinglitter homogenization, pregrinding (rather than clipping) aids in subsamplingof the decomposed litter for downstream analyses. Homogenized litter (5 g) wasplaced into nylon membrane bags with 0.22-μm pores (cat. no. SPEC17970; TischScientific). This pore size was selected to allow for the movement of water andnutrients but prevent the dispersal of exogenous microbial cells into the bags.The litterbags were then sterilized with at least 22 kGy gamma irradiation(Sterigenics). Microbial growth was not observed when the irradiated litter wasplated on agar media, but we recognize that complete sterilization is unlikely.However, our goal was to knock down the existing community to such lowabundance that the inocula communities could become established, which isconfirmed by our results. To create the five microbial inocula, four samples oflitter from each of the sites was collected on September 11, 2015, ground,homogenized within site, and 50 mg inoculum was added to each sterile litterbagto manipulate microbial community origin.
We placed 60litterbags in each site such that four replicates of each microbial communitytreatment could be collected every 6 mo for 18 mo (5 sites × 5 inocula × 4replicate plots × 3 time points = 300 litterbags). Replicates were distributedacross four 1 m × 1 m plots separated by >5 m. Litterbags from each plotwere collected on April 5, 2016, October 24, 2016, and on April 18, 2017. Toassess the composition of the in situ microbial communities outside thelitterbags, litter adjacent to the plots (the mixture of decaying plant speciespresent) was collected at initial deployment and at each litterbag collection(5 sites × 4 replicate plots × 4 time points = 80 in situ survey samples).
To compare thedecomposition rate within the transplant bags versus in situ litterdecomposition, we deployed two additional sets of litterbags within each plot(2 types × 5 sites × 4 plots × 3 time points = 120 litterbags). One set of bagscontained local, clipped (not ground) litter placed in litterbags made of 2-mmmesh window screening (in situ open). To test how the nylon membrane meshcontributed to these differences, the second set contained local, clippedlitter placed in the nylon membrane litterbags (in situ closed).
For information on howtemperature, precipitation, decomposition, litter chemistry, and bacterial andfungal abundance were assessed, and full details on sample processing, DNAextraction, genetic analysis, and bioinformatics, please seeSI Appendix,SupplementalMaterials and Methods. Briefly, bacterialcommunity composition was characterized using the V4 region of the 16Sribosomal RNA gene and fungal composition was characterized using the ITS2region with Illumina MiSeq. Sequences were submitted to the National Center forBiotechnology Information Sequence Read Archive under accession no. SRP150375.All bioinformatics processing was conducted in UPARSEversion 10. Analyses wereconducted by defining both 97% OTUs and exact sequence variants, but since theresults were nearly identical, we only present the analyses using 97% OTUs.
Results: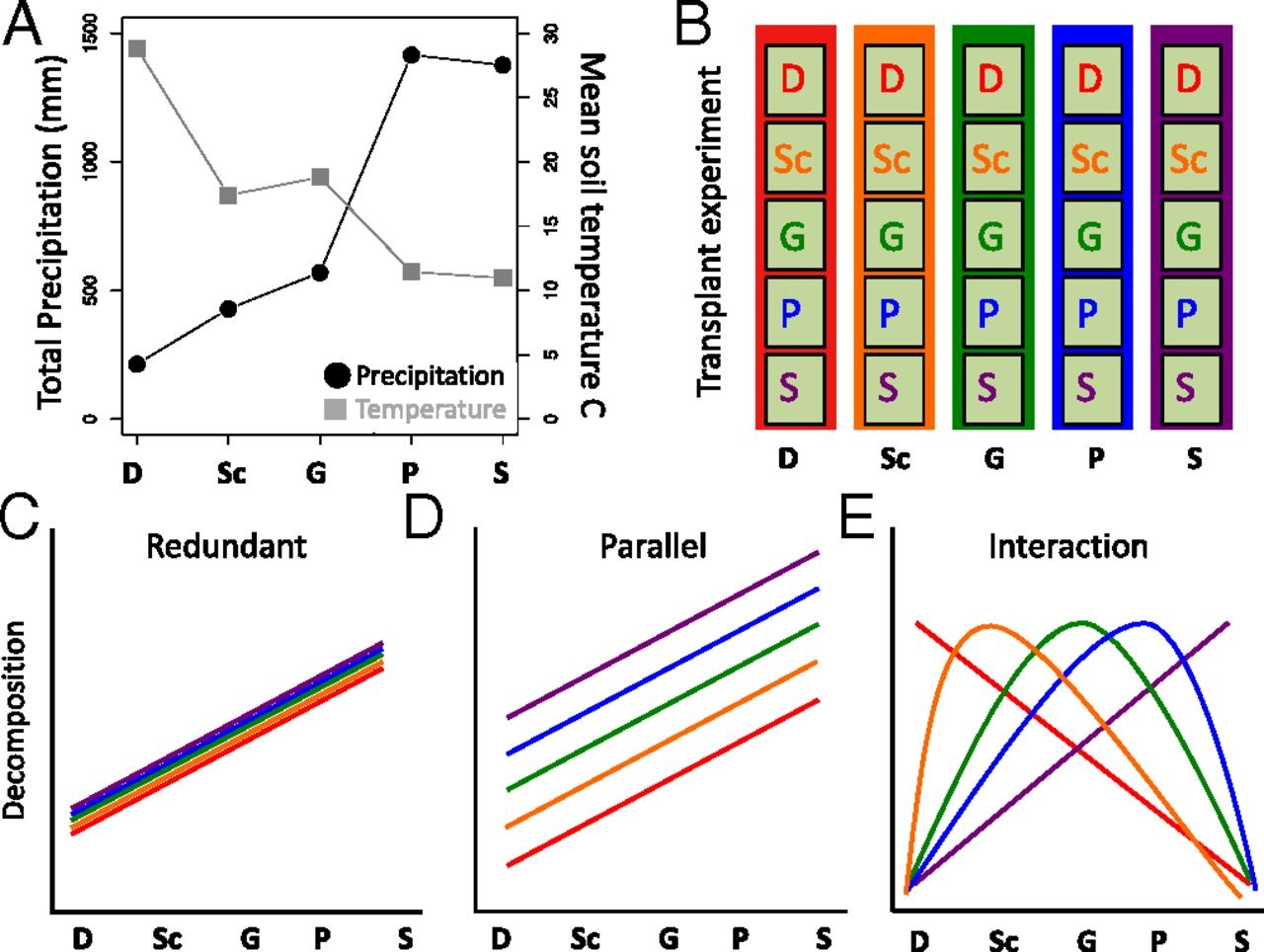
Fig. 1.
(A) Totalprecipitation (in millimeters) and mean annual soil temperature (in degreesCelsius) at the five sites along the elevation gradient. Sites are representedin increasing precipitation order in the same color scheme: desert (D) = red,scrubland (Sc) = orange, grassland (G) = green, pine–oak (P) = blue, andsubalpine (S) = purple. (B) Schematic of microbial transplantexperiment. Microbial communities from all sites were placed in a common gardenexperiment in all sites using a common substrate (irradiated grassland litterrepresented with light green box; n = 5 inocula × 5 sites × 4plots × 3 time points = 300 litterbags). Three possibilities for decompositionresponses are: (C) redundant, in which all microbes function similarlyin every site and are only affected by abiotic conditions; (D) parallel,in which microbes differentially affect decomposition, but respond to climatein a proportional manner; and (E) and interaction, in whichdecomposition is a result of an interaction between microbial communities andtheir environment. While any interaction is possible, we illustrate an examplein which a community decomposes most in its home site (home-field advantage).
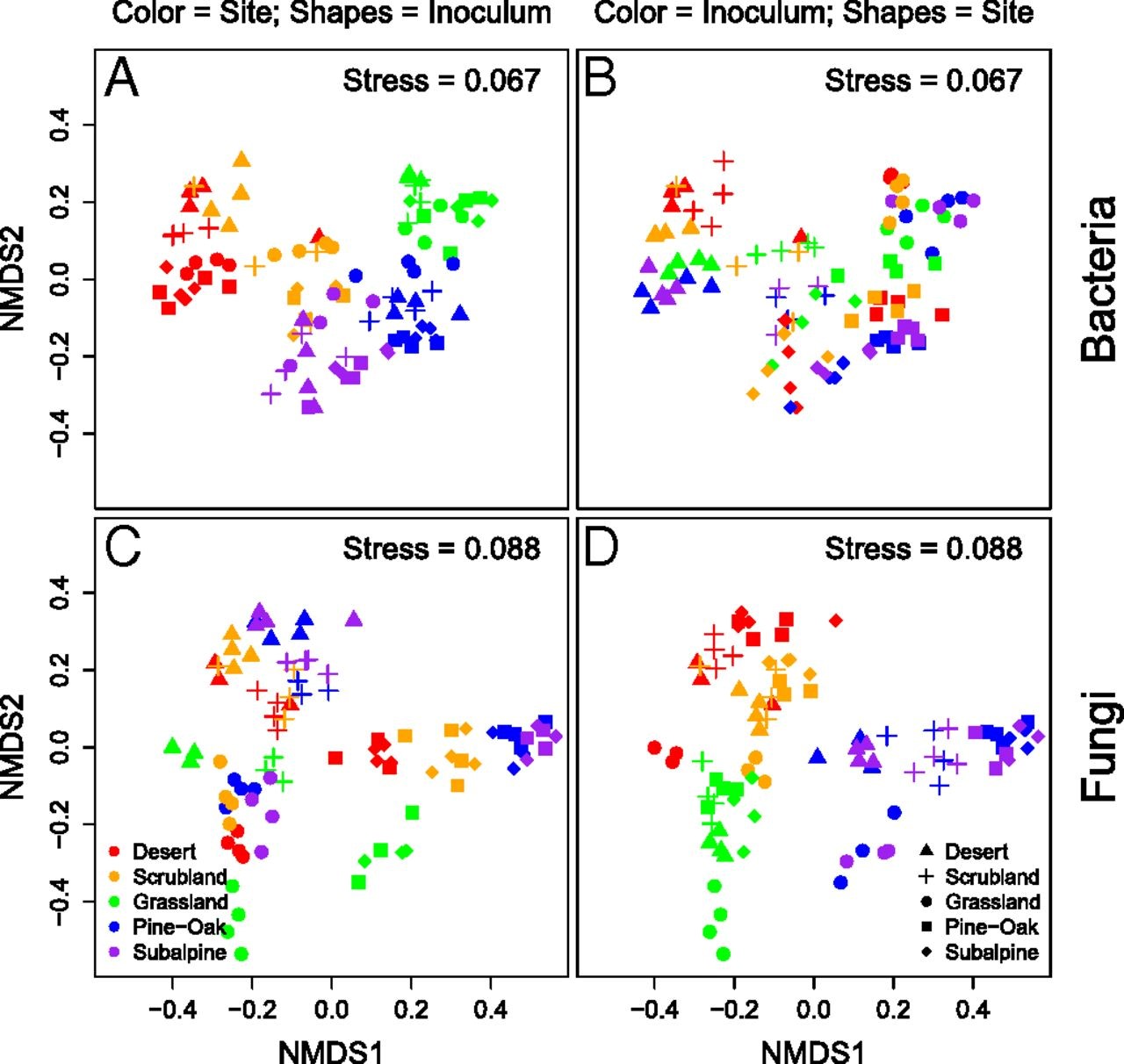
Fig. 2.
NMDS ofBray–Curtis microbial community composition at 18 mo for (A) bacteriacolored by site and shapes by inoculum and (B) bacteria colored byinoculum and shapes by site. The Bottom two panels are bothfungal community composition with either (C) colored by site and shapesby inoculum or (D) colored by inoculum and shapes by site.
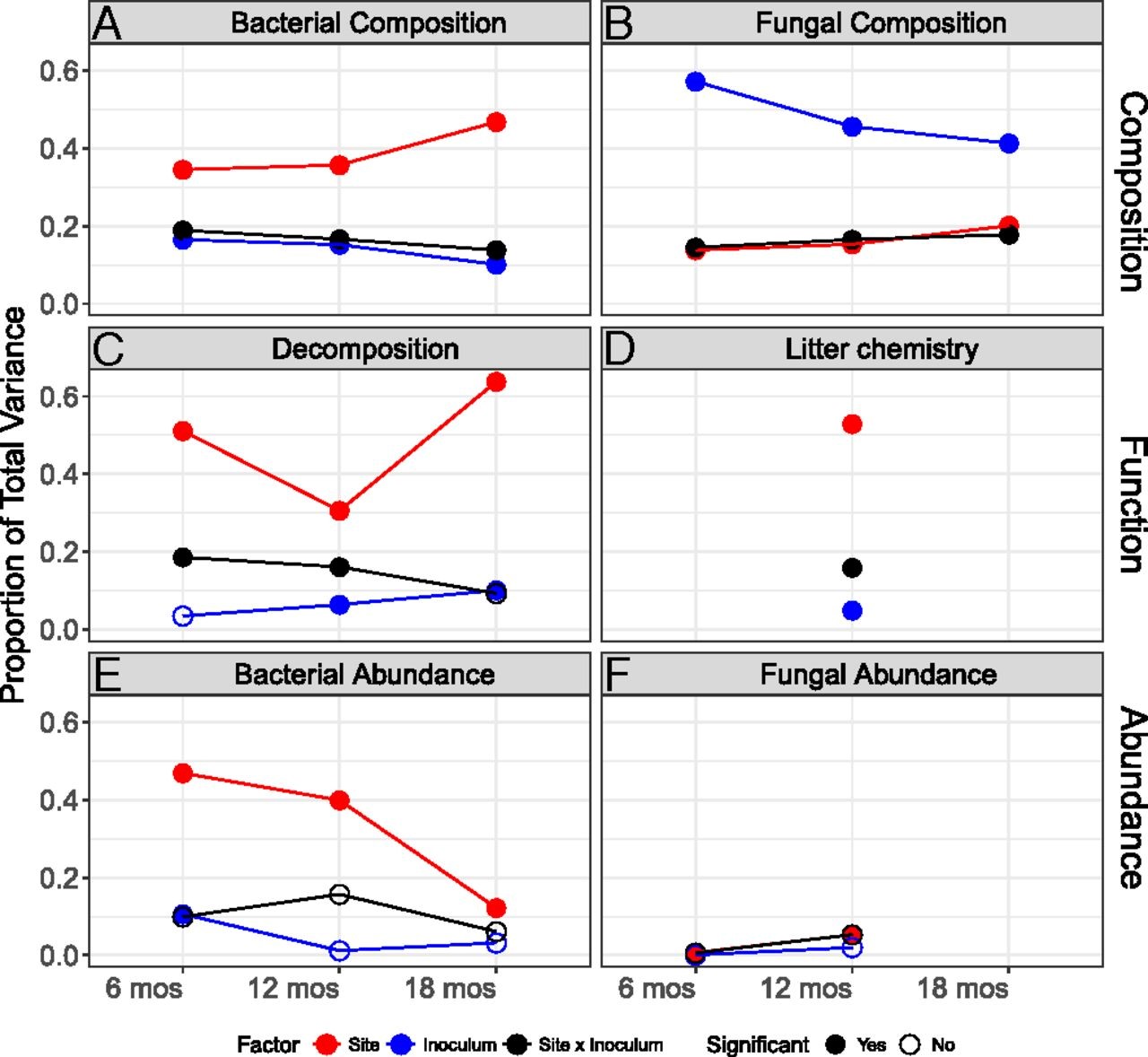
Fig. 3.
Proportion ofvariance explained by the treatments (site, inoculum, site x inoculum) on (A)bacterial community composition, (B) fungal community composition, (C)decomposition, (D) litter chemistry, (E) bacterial abundance, and(F) fungal abundance. The proportions for bacterial and fungal communitycomposition and litter chemistry are calculated based on variance estimatesfrom PERMANOVA (SI Appendix, Tables S4, S5, and S7), whereas those for microbial abundanceand decomposition are calculated from the total variance explained by two-wayANOVA multiplied by the partial eta-squares for each explanatory variable (SI Appendix, Tables S6 and S9).
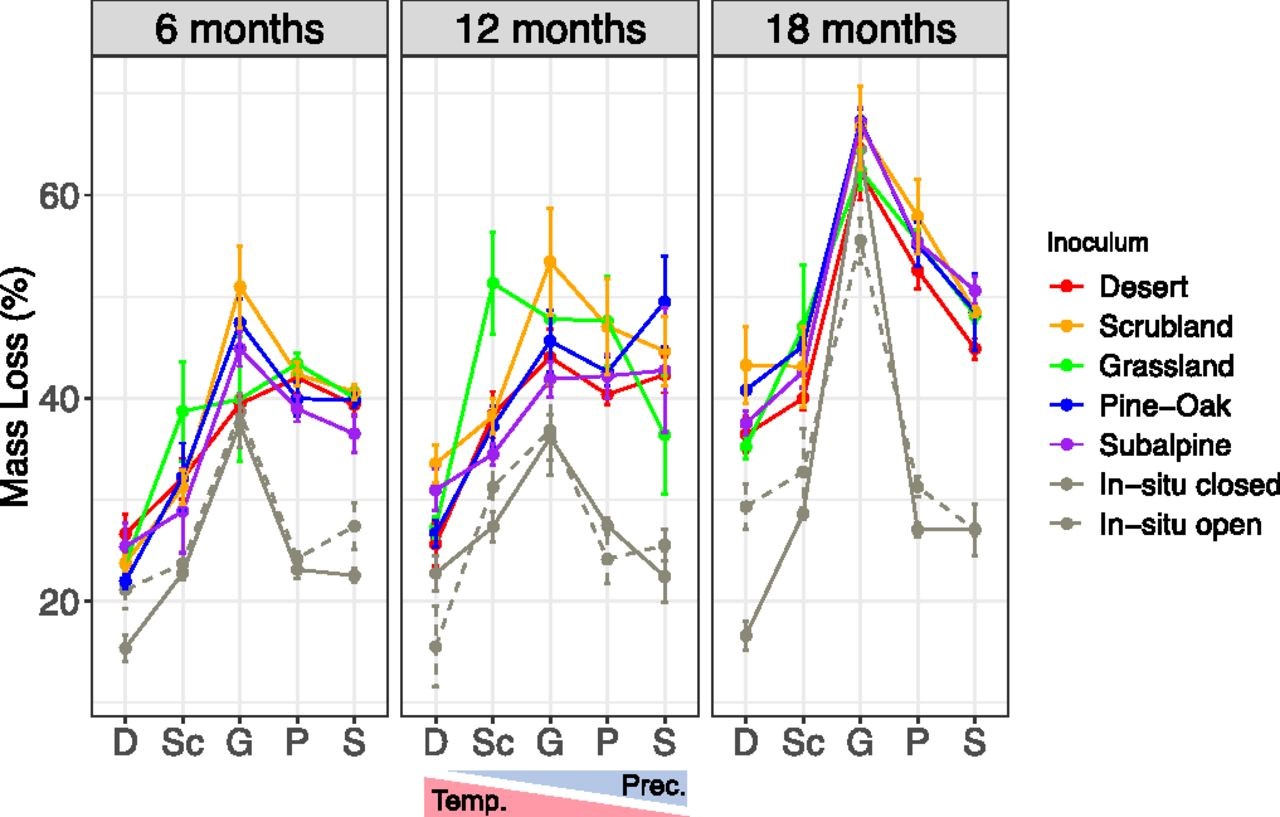
Fig. 4.
Variation inleaf litter decomposition (mean ± SE percent mass loss) for the full factorialtransplant experiment (5 inoculum treatments × 5 sites) along the gradientacross the three time points. Sites ordered in order of increasingprecipitation: D, desert; G, grassland; P, pine–oak; S, subalpine; and Sc,scrubland. In addition to transplant litterbags, we included open or closed insitu litterbags for comparison.
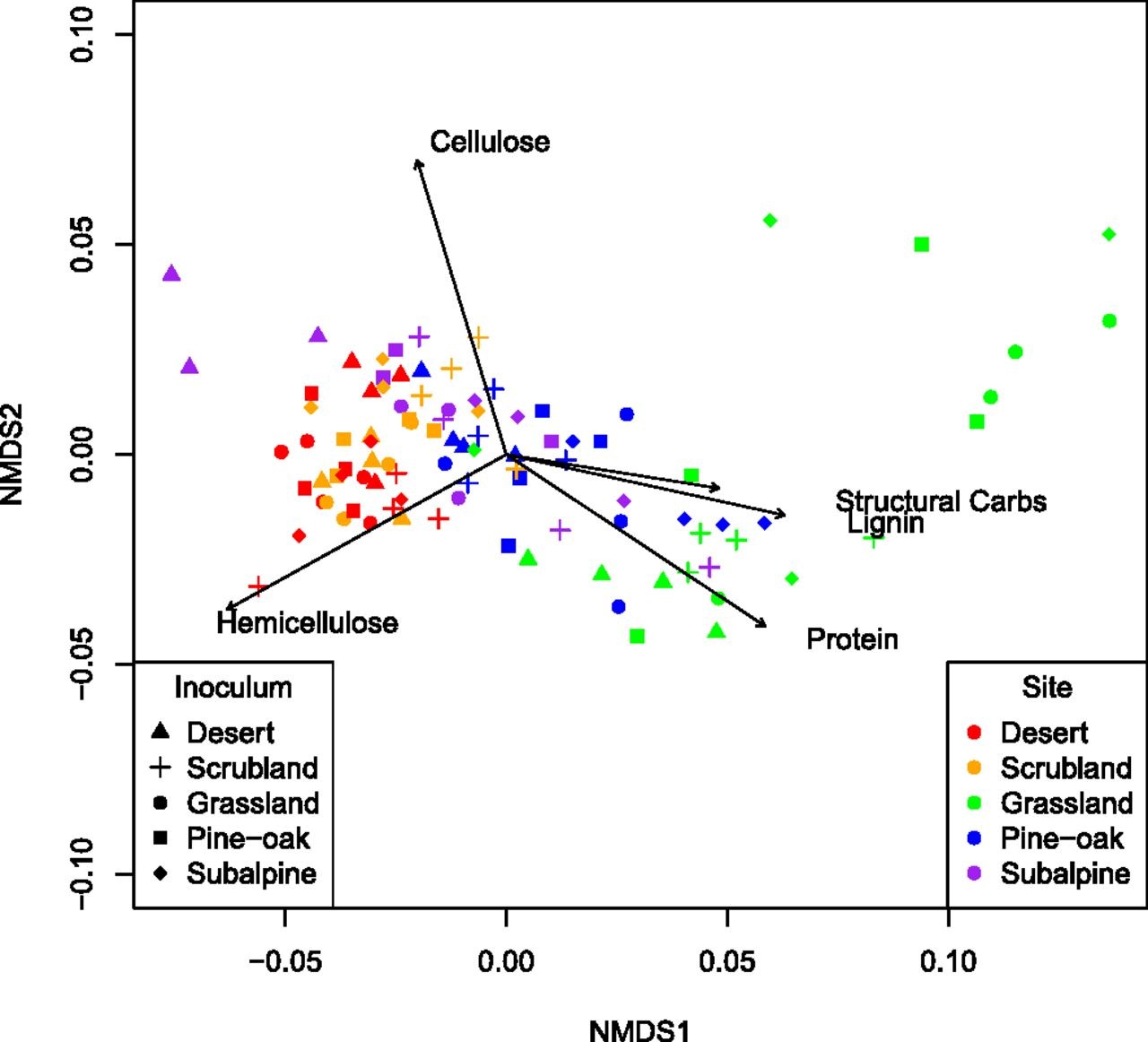
Fig. 5.
NMDS of litterchemistry of transplanted litterbags at 12 mo. Each point represents thechemical composition of the litter from each litterbag for each site (color)and inoculum (shape) combination (4 plots × 4 inocula × 5 sites = 100). Eachvector represents whether each of the five organic compounds (cellulose, crudeproteins, hemicellulose, lignin, and structural carbohydrates) increases ordecreases in abundance in that site. Stress = 0.045.
Discussion:
Experimentalmanipulations of abiotic and biotic factors are essential for disentangling themechanisms through which climate change will affect biodiversity and ecosystemprocesses. By performing a fully reciprocal microbial transplant experimentacross a gradient of nearly 2,000-m elevation and 15 °C soil temperature, wewere able to expand upon previous findings from laboratory manipulations thatshow that microbial composition can influence decomposition. We find thatnatural communities of microbial decomposers are not only functionallydistinct, but their functioning depends on an interaction between the communityand its climate, supporting a functional interaction model (Fig. 1E). Moreover,community differences accounted for a large fraction of the variation indecomposition observed. After a year, the abiotic environment was still thelargest factor explaining decomposition rates (30%) and litter chemistry (53%),but the microbial community and its interactions with the environment alsoaccounted for 22% of the variation in decomposition rate and 21% of thevariation in litter chemistry. These results counter the often implicitassumption that the contribution of microbial community composition todecomposition is negligible relative to climate or litter quality.
Whiledecomposition responses to climate depended on the microbial community, theywere not consistent with home-field advantage. Surprisingly, microbialcommunities from the extreme sites along the gradient were functionally similaracross the climate gradient, with desert microbes decomposing just as muchlitter in the subalpine environment as subalpine microbes and vice versa. Atthe same time, communities from the midelevation sites appeared to performopposite from what would be predicted by home-field advantage, with thescrubland microbes performing best in grassland and the grassland microbesperforming best in scrubland, with decomposition rates varying by as much as40% in a single environment (Fig. 4). Notably, home-field advantage is usuallyconsidered in terms of litter quality, whereas in this experiment we keptlitter quality constant and considered performance in terms of a home climate.Nonetheless, other studies have found inconsistent effects of home-field advantageor observed that these effects were limited to recalcitrant litter types.Ultimately, although the exact mechanisms remain unclear, our results suggestthat ecosystem predictions can be improved by considering the relationshipbetween a microbial community’s ability to degrade leaf litter and itsresponse to new climate conditions.
In addition tohaving large impacts on decomposition rates, the microbial community also hadsmaller but significant effects on the types of carbon compounds degraded. Thisis important because the types of carbon compounds left behind can influencecarbon storage. These results indicate that litter communities from differentenvironments vary in carbohydrate degradation traits, similar to previous workin the grassland site, showing that communities subjected to drought conditionsshift in composition and glycoside hydrolase (GH) abundance.
The duration ofour experiment also allowed us to tease apart the timing of the bacterial andfungal compositional shifts and their corresponding functional consequences.After 18 mo, bacterial composition was more reflective of their new environmentthan of their initial inoculum, while fungal composition still primarilyreflected the initial inoculum. This result—that fungal communities are moreresistant to change—was also found in an earlier litter transplant experimentwithin the grassland site. One potential explanation is faster rates ofturnover in bacterial communities, which are known to respond more quickly todisturbancesthan fungi.
The timing ofthese shifts relative to the functional consequences of the inoculatedcommunity suggests that bacterial decomposers may be less functionallyredundant than fungal decomposers and that they might have larger effects ondecomposition than previously believed. The effect of the microbial community(and particularly the community-by-site interaction) largely attenuated from 12to 18 mo, even though the fungal communities within a site were still highlydistinct at 18 mo. A review of the subject also concluded that shifts in fungalcommunity structure do not necessarily influence decomposition rates, potentiallydue to the high overlap in metabolic activities of saprobic soil fungal species.Indeed, both aquatic and terrestrial laboratory manipulations found that fungaldiversity–decomposition relationships saturated rapidly after the addition ofonly two to six species. In contrast, a study that manipulated bacterialrichness found increasing respiration function with diversity beyond 72species. These results might be attributed to the larger breadth ofphylogenetic diversity and thus corresponding functional traits represented bythe bacterial communities. In our study, >95% of fungal taxa belonged to asingle phylum (Ascomycota), whereas the vast majority of bacteria belonged tofour phyla, representing hundreds of millions of years of evolution between them.In addition, a large diversity of GH genes have been found in bacteria fromleaf litter and soil. Interestingly, recent work on soil microbial decomposerssuggests that both fungi and bacteria are involved in complex organic matterbreakdown, and that interactions between fungi and bacteria in decompositionare perhaps more lateral and less hierarchical than previously believed. Thus,it is becoming clear that bacteria can have strong impacts on decomposition.The larger phylogenetic diversity and breadth of metabolic capacities ofbacteria may explain why shifts in bacterial composition appear to have astronger effect on decomposition than shifts in fungal diversity.
Finally, it isnotable that microbial composition, but not initial microbial biomass,predicted litter decomposition rates. Decomposition within a site was notcorrelated with fungal or bacterial biomass in the inoculum leaf litter. Forinstance, the desert community, with its low initial inoculum biomass, carriedout decomposition at similar rates to communities with greater biomass. At thesame time, microbial biomass and decomposition were positively correlatedacross all samples and sites. Many studies have observed similar correlations;for instance, a recent study found that soil microbial biomass, as measured bysubstrate-induced respiration, was correlated with leaf litter decomposition asmuch as litter quality and climate. However, our transplant experimentindicates that this correlation is not due to microbial biomass driving decomposition.Instead, biomass and decomposition might be correlated because they are bothinfluenced by environment. Alternatively, faster decomposition might result inhigher microbial biomass.
In conclusion,decomposition responses to changing temperature and precipitation depended onthe composition of a microbial decomposer community. This study examines thedecomposition response curves of different microbial communities across a rangeof climate conditions that could be relevant for predictions of ecosystemfunctioning. In fact, the dominant plant taxa along this same elevationgradient have shifted upward over the last 30 y due to climate change. While itis impossible to know if the microbial communities have also begun to shifttheir range due to a lack of historical data, our study indicates thatbacterial and fungal decomposer communities take time to respond to changes inclimate, and this lag has consequences for functioning. Future work should alsoconsider invertebrate grazers, viruses, and fungal–bacterial interactions toobtain a more complete understanding of decomposition responses to climatechange.